ศูนย์กลางที่แอคทีฟของโปรตีนและปฏิสัมพันธ์กับลิแกนด์ การทำงานของโปรตีน
คุณสมบัติหลักของโปรตีนที่ช่วยให้มั่นใจว่าหน้าที่ของมันคือปฏิสัมพันธ์แบบคัดเลือกกับสารบางชนิด - ลิแกนด์
ลิแกนด์สามารถเป็นสารที่มีลักษณะแตกต่างกันได้ทั้งสารประกอบที่มีน้ำหนักโมเลกุลต่ำและโมเลกุลขนาดใหญ่รวมถึงโปรตีน ในโมเลกุลของโปรตีนมีบริเวณที่ยึดแกนด์ - ศูนย์จับหรือศูนย์ที่ใช้งานอยู่ ศูนย์การจับถูกสร้างขึ้นจากเรซิดิวของกรดอะมิโนที่นำมารวมกันอันเป็นผลมาจากการก่อตัวของโครงสร้างทุติยภูมิและตติยภูมิ
พันธะระหว่างโปรตีนและลิแกนด์สามารถเป็นแบบไม่มีโควาเลนต์หรือโควาเลนต์ได้ ความจำเพาะสูงของปฏิสัมพันธ์ ("การรับรู้") ของโปรตีนและลิแกนด์นั้นมาจากการเสริมกันของโครงสร้างของศูนย์กลางการจับกับโครงสร้างเชิงพื้นที่ของลิแกนด์
ความสอดคล้องกันเป็นที่เข้าใจกันว่ามีความสอดคล้องกันทางเคมีและเชิงพื้นที่ระหว่างศูนย์กลางที่แอคทีฟของโปรตีนและลิแกนด์ ปฏิกิริยาระหว่างโปรตีน P และลิแกนด์ L อธิบายโดยสมการ:
โปรตีน + ลิแกนด์↔ โปรตีนลิแกนด์คอมเพล็กซ์
1. คุณสมบัติทางเคมีกายภาพหลักของโปรตีนคือน้ำหนักโมเลกุล ประจุไฟฟ้า และความสามารถในการละลายในน้ำ น้ำหนักโมเลกุลของโปรตีนอาจแตกต่างกันมาก ตัวอย่างเช่น ฮอร์โมนอินซูลินมีน้ำหนักโมเลกุลประมาณ 6,000 Da ในขณะที่อิมมูโนโกลบูลิน M มีน้ำหนักโมเลกุลประมาณ 1 ล้านดา น้ำหนักโมเลกุลของโปรตีนขึ้นอยู่กับจำนวนของกรดอะมิโนที่ตกค้างที่ประกอบเป็นองค์ประกอบ เช่นเดียวกับมวลของส่วนประกอบที่ไม่ใช่กรดอะมิโน มวลของกรดอะมิโน 1 ตัวมีค่าเฉลี่ย 110 Da ดังนั้น เมื่อทราบจำนวนกรดอะมิโนตกค้างในโปรตีน เราจึงสามารถประมาณน้ำหนักโมเลกุลของโปรตีนได้ และในทางกลับกัน (N.N. Mushkambarov, 1995) ประจุไฟฟ้าของโปรตีนถูกกำหนดโดยอัตราส่วนของกลุ่มที่มีประจุบวกและลบบนพื้นผิวของโมเลกุล ประจุของอนุภาคโปรตีนขึ้นอยู่กับ pH ของตัวกลาง แนวคิดของ "จุดไอโซอิเล็กทริก" ใช้เพื่อกำหนดลักษณะเฉพาะของโปรตีน จุดไอโซอิเล็กทริก (pI) - ค่า pH ของตัวกลางที่ประจุทั้งหมดของอนุภาคโปรตีน ศูนย์. ที่จุดไอโซอิเล็กทริก โปรตีนมีความคงตัวน้อยที่สุดในสารละลายและตกตะกอนได้ง่าย ค่า pI ขึ้นอยู่กับอัตราส่วนของกรดอะมิโนที่เป็นกรดและด่างในโปรตีน สำหรับโปรตีนและเปปไทด์ที่มีกรดอะมิโนที่เป็นกรดมากกว่า (ประจุลบที่ pH 7.0) ค่า pI จะอยู่ในสภาพแวดล้อมที่เป็นกรด สำหรับโปรตีนและเปปไทด์ที่มีความโดดเด่นของกรดอะมิโนพื้นฐาน (ประจุบวกที่ pH 7.0) ค่า pI จะอยู่ในสภาพแวดล้อมที่เป็นกรด จุดไอโซอิเล็กทริกเป็นค่าคงตัวของโปรตีน ค่าของมันสำหรับโปรตีนในเนื้อเยื่อสัตว์ส่วนใหญ่อยู่ในช่วง 5.5 ถึง 7.0 ซึ่งบ่งชี้ถึงความเด่นของกรดอะมิโนที่เป็นกรดในองค์ประกอบ อย่างไรก็ตามในธรรมชาติมีโปรตีนที่มีค่าจุดไอโซอิเล็กทริกอยู่ที่ค่า pH สูงสุดของตัวกลาง โดยเฉพาะค่า pI ของเปปซิน (เอนไซม์น้ำย่อย) คือ 1 และค่าของไลโซไซม์ (เอ็นไซม์ที่ทำลายผนังเซลล์ของจุลินทรีย์) อยู่ที่ประมาณ 11 ค่าน้ำหนักโมเลกุลและค่าจุดไอโซอิเล็กทริกของโปรตีนบางชนิดแสดงให้เห็น ในตารางที่ 1.4 ตารางที่ 1.4 ค่าคงที่ของพลาสมาเลือดและโปรตีนเนื้อเยื่อ
ความสามารถในการละลายของโปรตีนในน้ำจากหลักสูตรเคมีชีวฟิสิกส์ เป็นที่ทราบกันว่าโปรตีนในฐานะสารประกอบโมเลกุลสูงจะก่อตัวเป็นสารละลายคอลลอยด์ ความคงตัวของสารละลายโปรตีนในน้ำถูกกำหนดโดยปัจจัยต่อไปนี้:
โปรดทราบว่าภายใต้อิทธิพลของปัจจัยทางกายภาพและทางเคมีต่างๆ การตกตะกอนของโปรตีนจากสารละลายคอลลอยด์สามารถเกิดขึ้นได้ แยกแยะ:
โปรดทราบว่ากลไกต่อไปนี้อาจรองรับปฏิกิริยาการตกตะกอนของโปรตีน:
ส่วนใหญ่แล้ว การกระทำของปัจจัยที่ก่อให้เกิดการตกตะกอนของโปรตีนนั้นมีลักษณะเฉพาะด้วยกลไกสองหรือทั้งสามอย่างรวมกัน กิจกรรมทางชีวภาพ การทำงานของโปรตีนใด ๆ ขึ้นอยู่กับความสามารถในการโต้ตอบกับโมเลกุลหรือไอออน - ลิแกนด์ที่กำหนดไว้อย่างเคร่งครัด ตัวอย่างเช่น สำหรับเอนไซม์ที่กระตุ้นปฏิกิริยาเคมี ลิแกนด์จะเป็นสารที่เกี่ยวข้องกับปฏิกิริยาเหล่านี้ (สารตั้งต้น) เช่นเดียวกับโคแฟคเตอร์ ตัวกระตุ้น และสารยับยั้ง สำหรับโปรตีนในการขนส่ง ลิแกนด์คือสารที่ขนส่ง เป็นต้น ลิแกนด์สามารถโต้ตอบกับตำแหน่งเฉพาะของโมเลกุลโปรตีน - ศูนย์กลางการจับหรือศูนย์แอคทีฟ ศูนย์นี้ถูกสร้างขึ้นโดยอนุมูลของกรดอะมิโนที่อยู่ใกล้เคียงกันที่ระดับของโครงสร้างระดับอุดมศึกษาของโปรตีน ความสามารถของแกนด์ในการโต้ตอบกับศูนย์รวมนั้นเกิดจากการเสริมกันนั่นคือความสมบูรณ์ร่วมกันของโครงสร้างเชิงพื้นที่ของพวกเขา (คล้ายกับปฏิสัมพันธ์ "กุญแจ - ล็อค") พันธะที่ไม่ใช่โควาเลนต์ (ไฮโดรเจน, อิออน, ไม่ชอบน้ำ) เกิดขึ้นระหว่างกลุ่มการทำงานของลิแกนด์และตำแหน่งการจับ ความสมบูรณ์ของลิแกนด์และตำแหน่งการจับสามารถอธิบายความจำเพาะสูง (คุณสมบัติการคัดเลือก) ของปฏิกิริยาระหว่างโปรตีนกับลิแกนด์ ดังนั้นโปรตีนที่แตกต่างกันจึงแตกต่างกันในคุณสมบัติทางเคมีกายภาพและฤทธิ์ทางชีวภาพ วิธีการแยกโปรตีนผสมออกเป็นเศษส่วนและแยกโปรตีนเอนไซม์แต่ละตัวขึ้นอยู่กับความแตกต่างเหล่านี้ วิธีการเหล่านี้ใช้กันอย่างแพร่หลายในด้านชีวเคมีทางการแพทย์และเทคโนโลยีชีวภาพ 2. การเสื่อมสภาพของโปรตีน- นี่คือการเปลี่ยนแปลงในธรรมชาติ (ธรรมชาติ) ทางเคมีกายภาพและที่สำคัญที่สุดคือคุณสมบัติทางชีวภาพของโปรตีนเนื่องจากการละเมิดโครงสร้างควอเทอร์นารีตติยภูมิและแม้กระทั่งทุติยภูมิ การเสื่อมสภาพของโปรตีนอาจเกิดจาก:
โปรตีนที่ทำให้เสียสภาพมีลักษณะโดย:
โปรดทราบว่าภายใต้เงื่อนไขบางประการ เป็นไปได้ที่จะฟื้นฟูโครงสร้างโปรตีน (ดั้งเดิม) ดั้งเดิมหลังจากการกำจัดปัจจัยที่ทำให้เกิดการเสียสภาพ กระบวนการนี้เรียกว่าการฟื้นฟู จำตัวอย่างการใช้กระบวนการทำให้โปรตีนเสื่อมสภาพในยา:
|
4(1). เฮโมโกลบินเป็นโปรตีนอัลโลสเตอริก การเปลี่ยนแปลงโครงสร้างในโมเลกุลของเฮโมโกลบิน ผลความร่วมมือ สารควบคุมความสัมพันธ์ของเฮโมโกลบินสำหรับออกซิเจน ความแตกต่างของโครงสร้างและหน้าที่ระหว่าง myoglobin และ hemoglobin
เฮโมโกลบิน: โปรตีนอัลโลสเตอริก
การเปลี่ยนแปลงเชิงวิวัฒนาการจากโมโนเมอร์ไมโอโกลบินไปเป็นเตตราเมอร์เฮโมโกลบินมาพร้อมกับคุณสมบัติใหม่ โมเลกุลของเฮโมโกลบินนั้นซับซ้อนกว่าโมเลกุลของไมโอโกลบินมาก ก่อนอื่นฮีโมโกลบินนอกเหนือจาก 0 2 ขนส่ง H + และ CO 2 ประการที่สอง การผูกมัดของออกซิเจนโดยเฮโมโกลบินถูกควบคุมโดยส่วนประกอบเฉพาะของสภาพแวดล้อมภายใน ได้แก่ H + , CO 2 และสารประกอบอินทรีย์ฟอสฟอรัส สารควบคุมเหล่านี้มีอิทธิพลอย่างมากต่อความสามารถของฮีโมโกลบินในการจับออกซิเจน แม้ว่าจะเกาะติดกับโปรตีนในบริเวณที่ห่างไกลจากฮีมก็ตาม โดยทั่วไป สิ่งที่เรียกว่า ปฏิสัมพันธ์ allosteric,เหล่านั้น. ปฏิสัมพันธ์ระหว่างพื้นที่ที่แยกจากกันเกิดขึ้นในโปรตีนหลายชนิด เล่นเอฟเฟกต์ allosteric บทบาทสำคัญในการควบคุมและบูรณาการกระบวนการระดับโมเลกุลในระบบชีวภาพ เฮโมโกลบินเป็นโปรตีน allosteric ที่มีการศึกษามากที่สุด ดังนั้นจึงควรคำนึงถึงโครงสร้างและหน้าที่ในรายละเอียดเพิ่มเติม
การเปลี่ยนแปลงทางโครงสร้างในฮีโมโกลบิน
การผูกมัดของออกซิเจนมาพร้อมกับการแตกของเกลือ
พันธะที่เกิดจากหมู่เทอร์มินอลคาร์บอกซิล
หน่วยย่อย (รูปที่ 7) สิ่งนี้อำนวยความสะดวกในการจับกันของโมเลกุลต่อไปนี้
ออกซิเจนเพราะมันต้องการการแตกของจำนวนที่น้อยกว่า
พันธะเกลือ การเปลี่ยนแปลงเหล่านี้มีผลกระทบอย่างมากต่อ
โครงสร้างทุติยภูมิ ตติยภูมิ และควอเทอร์นารีโดยเฉพาะอย่างยิ่ง
เฮโมโกลบิน. ในกรณีนี้ หน่วยย่อย A / B หนึ่งคู่จะเปลี่ยน
เทียบกับคู่ A/B อื่น ซึ่งนำไปสู่การกระชับ
tetramer และความสัมพันธ์ของ heme ที่เพิ่มขึ้นสำหรับออกซิเจน (รูปที่ 8 และ 9)
การเปลี่ยนแปลงเชิงโครงสร้างในสภาพแวดล้อมของฮีโมกรุ๊ป
ออกซิเจนของเฮโมโกลบินมาพร้อมกับโครงสร้าง
การเปลี่ยนแปลงในสภาพแวดล้อมของเฮโมกรุ๊ป เมื่อให้ออกซิเจน อะตอม
เหล็กซึ่งใน deoxyhemoglobin ยื่นออกมา 0.06 นาโนเมตรจาก
ระนาบของวงแหวน heme ดึงเข้าไปในระนาบนี้ (รูปที่
สิบ). ตามอะตอมของเหล็ก มันจะเคลื่อนเข้าใกล้ฮีมมากขึ้น
ฮิสติดีนส่วนต้น (F8) เช่นเดียวกับเพื่อนบ้านที่เกี่ยวข้อง
โมเลกุลของเฮโมโกลบินสามารถอยู่ในสองรูปแบบ - ตึงเครียดและผ่อนคลาย รูปแบบที่ผ่อนคลายของเฮโมโกลบินมีแนวโน้มที่จะอิ่มตัวด้วยออกซิเจนเร็วกว่าออกซิเจน 70 เท่า การเปลี่ยนแปลงในเศษส่วนของรูปแบบความตึงเครียดและผ่อนคลายในปริมาณทั้งหมดของฮีโมโกลบินในเลือดกำหนดรูปแบบ S-shape ของเส้นโค้งการแยกตัวของออกซีเฮโมโกลบินและด้วยเหตุนี้จึงเรียกว่าความสัมพันธ์ของฮีโมโกลบินสำหรับออกซิเจน หากความน่าจะเป็นของการเปลี่ยนจากเฮโมโกลบินในรูปแบบตึงเครียดไปเป็นแบบผ่อนคลายมากขึ้น ความสัมพันธ์ของฮีโมโกลบินต่อออกซิเจนจะเพิ่มขึ้น และในทางกลับกัน ความน่าจะเป็นของการก่อตัวของเศษส่วนของเฮโมโกลบินเหล่านี้เปลี่ยนแปลงขึ้นหรือลงภายใต้อิทธิพลของปัจจัยหลายประการ ปัจจัยหลักคือการผูกมัดของออกซิเจนกับกลุ่มฮีมของโมเลกุลเฮโมโกลบิน ยิ่งไปกว่านั้น ยิ่งกลุ่มเฮโมโกลบินของเฮโมโกลบินจับออกซิเจนในเม็ดเลือดแดงมากเท่าไร โมเลกุลของเฮโมโกลบินก็จะเปลี่ยนไปอยู่ในรูปแบบที่ผ่อนคลายได้ง่ายขึ้นเท่านั้น และความสัมพันธ์ของพวกมันกับออกซิเจนก็จะสูงขึ้น ดังนั้น ที่ระดับ P02 ต่ำ ซึ่งเกิดขึ้นในเนื้อเยื่อที่มีการเผาผลาญ ความสัมพันธ์ของฮีโมโกลบินกับออกซิเจนจึงต่ำกว่า และที่ P02 สูงก็จะยิ่งสูงขึ้น ทันทีที่เฮโมโกลบินจับออกซิเจน ความสัมพันธ์กับออกซิเจนจะเพิ่มขึ้น และโมเลกุลของเฮโมโกลบินจะอิ่มตัวเมื่อจับกับโมเลกุลออกซิเจนสี่ตัว เมื่อเซลล์เม็ดเลือดแดงที่มีเฮโมโกลบินไปถึงเนื้อเยื่อ ออกซิเจนจากเซลล์เม็ดเลือดแดงจะกระจายเข้าสู่เซลล์ ในกล้ามเนื้อจะเข้าสู่คลังออกซิเจนชนิดหนึ่ง - เข้าไปในโมเลกุลของ myoglobin ซึ่งใช้ออกซิเจนในการออกซิเดชันทางชีวภาพของกล้ามเนื้อ การแพร่กระจายของออกซิเจนจากเม็ดเลือดแดงเฮโมโกลบินไปยังเนื้อเยื่อเกิดจาก P02 ต่ำในเนื้อเยื่อ - 35 มม. ปรอท ศิลปะ. ภายในเซลล์เนื้อเยื่อ ความตึงเครียดของออกซิเจนที่จำเป็นต่อการรักษาระดับเมตาบอลิซึมตามปกติจะเล็กลงอีก - ไม่เกิน 1 kPa ดังนั้นออกซิเจนโดยการแพร่กระจายจากเส้นเลือดฝอยไปถึงเซลล์ที่มีการเผาผลาญ เนื้อเยื่อบางชนิดถูกปรับให้เข้ากับปริมาณ PO2 ต่ำในเส้นเลือดฝอย ซึ่งชดเชยด้วยความหนาแน่นสูงของเส้นเลือดฝอยต่อปริมาตรของเนื้อเยื่อ ตัวอย่างเช่น ในกล้ามเนื้อโครงร่างและหัวใจ เส้นเลือดฝอย P02 สามารถลดลงอย่างรวดเร็วอย่างมากในระหว่างการหดตัว เซลล์กล้ามเนื้อประกอบด้วยโปรตีน myoglobin ซึ่งมีความสัมพันธ์กับออกซิเจนสูงกว่าเฮโมโกลบิน Myoglobin อิ่มตัวด้วยออกซิเจนอย่างเข้มข้นและมีส่วนช่วยในการแพร่กระจายจากเลือดไปยังกล้ามเนื้อโครงร่างและกล้ามเนื้อหัวใจซึ่งจะกำหนดกระบวนการของการเกิดออกซิเดชันทางชีวภาพ เนื้อเยื่อเหล่านี้สามารถดึงออกซิเจนได้มากถึง 70% จากเลือดที่ไหลผ่าน ซึ่งเกิดจากการที่ความสัมพันธ์ของฮีโมโกลบินกับออกซิเจนลดลงภายใต้อิทธิพลของอุณหภูมิและค่า pH ของเนื้อเยื่อ ผลของ pH และอุณหภูมิต่อความสัมพันธ์ของเฮโมโกลบินต่อออกซิเจน โมเลกุลของเฮโมโกลบินสามารถทำปฏิกิริยากับไฮโดรเจนไอออนได้ อันเป็นผลมาจากปฏิกิริยานี้ ทำให้ความสัมพันธ์ของเฮโมโกลบินกับออกซิเจนลดลง เมื่อความอิ่มตัวของฮีโมโกลบินน้อยกว่า 100% ค่า pH ต่ำจะลดการจับของออกซิเจนกับเฮโมโกลบิน - เส้นโค้งการแยกตัวของออกซีเฮโมโกลบินจะเลื่อนไปทางขวาตามแนวแกน x การเปลี่ยนแปลงคุณสมบัติของเฮโมโกลบินภายใต้อิทธิพลของไฮโดรเจนไอออนนี้เรียกว่าปรากฏการณ์บอร์ เนื้อเยื่อที่ทำงานเมตาบอลิซึมจะผลิตกรด เช่น กรดแลคติกและคาร์บอนไดออกไซด์ หากค่า pH ของเลือดในพลาสมาลดลงจากปกติ 7.4 เป็น 7.2 ซึ่งเกิดขึ้นระหว่างการหดตัวของกล้ามเนื้อ ความเข้มข้นของออกซิเจนในพลาสมาจะเพิ่มขึ้นเนื่องจากผลของบอร์ ตัวอย่างเช่น ที่ pH คงที่ 7.4 เลือดจะให้ออกซิเจนประมาณ 45% กล่าวคือ ความอิ่มตัวของฮีโมโกลบินกับออกซิเจนจะลดลงเป็น 55% อย่างไรก็ตาม เมื่อ pH ลดลงเหลือ 7.2 เส้นโค้งการแยกตัวจะเลื่อนไปตามแกน x ไปทางขวา เป็นผลให้ความอิ่มตัวของออกซิเจนของเฮโมโกลบินลดลงถึง 40% กล่าวคือ เลือดสามารถให้ออกซิเจนในเนื้อเยื่อได้มากถึง 60% ซึ่งมากกว่าค่า pH คงที่ 1/3 เนื้อเยื่อที่ใช้งานเมตาบอลิซึมเพิ่มการผลิตความร้อน อุณหภูมิของเนื้อเยื่อที่เพิ่มขึ้นระหว่างการทำงานทางกายภาพจะเปลี่ยนอัตราส่วนของเศษส่วนของเฮโมโกลบินในเม็ดเลือดแดงและทำให้เส้นโค้งการแยกตัวของออกซีเฮโมโกลบินไปทางขวาตามแนวแกน x ผลที่ตามมา ปริมาณมากออกซิเจนจะถูกปล่อยออกจากฮีโมโกลบินของเม็ดเลือดแดงและเข้าสู่เนื้อเยื่อ ผลของ 2,3-diphosphoglycerate (2,3-DPG) ต่อความสัมพันธ์ของเฮโมโกลบินต่อออกซิเจน ภายใต้สภาวะทางสรีรวิทยาบางอย่าง เช่น เมื่อ P02 ในเลือดต่ำกว่าปกติ (ภาวะขาดออกซิเจน) อันเป็นผลมาจากบุคคลที่อยู่ในระดับความสูงเหนือระดับน้ำทะเล การจัดหาออกซิเจนไปยังเนื้อเยื่อจะไม่เพียงพอ ในระหว่างการขาดออกซิเจน ความสัมพันธ์ของฮีโมโกลบินกับออกซิเจนอาจลดลงเนื่องจากการเพิ่มขึ้นของเนื้อหาของ 2,3-DPG ในเม็ดเลือดแดง ในทางตรงกันข้ามกับผลกระทบของ Bohr การลดลงของความสัมพันธ์ของฮีโมโกลบินสำหรับออกซิเจนภายใต้อิทธิพลของ 2,3-DPG นั้นไม่สามารถย้อนกลับได้ในเส้นเลือดฝอยของปอด อย่างไรก็ตาม เมื่อเลือดเคลื่อนผ่านเส้นเลือดฝอยของปอด ผลของ 2,3-DPG ต่อการลดการก่อตัวของ oxyhemoglobin ในเม็ดเลือดแดง (ส่วนที่แบนของเส้นโค้งการแยกตัวของออกซีฮีโมโกลบิน) จะเด่นชัดน้อยกว่าการปล่อยออกซิเจนภายใต้อิทธิพลของ 2,3-DPG ในเนื้อเยื่อ (ส่วนที่ลาดเอียงของเส้นโค้ง) ซึ่งให้ออกซิเจนตามปกติไปยังเนื้อเยื่อ
โครงสร้างสามมิติดั้งเดิมถูกสร้างขึ้นจากการกระทำของพลังงานและปัจจัยเอนโทรปีจำนวนหนึ่ง การเปลี่ยนแปลงในสถานะโครงสร้างของโมเลกุลโปรตีนอันเนื่องมาจากอิทธิพลภายนอกต่างๆ (pH, อุณหภูมิ, องค์ประกอบไอออนิก) ก็สะท้อนให้เห็นในกิจกรรมการทำงานของมันเช่นกัน การจัดเรียงโครงสร้างใหม่เกิดขึ้นอย่างรวดเร็ว ในระยะแรก พวกมันมีลักษณะโครงสร้างจุลภาคในท้องถิ่น ทำให้เกิดการกระจัดของกลุ่มอะตอมแต่ละกลุ่มเท่านั้น การแพร่กระจายของการกระจัดในพื้นที่ดังกล่าวไปยังบริเวณอื่น ๆ ของโครงสร้างโมเลกุลขนาดใหญ่จะนำไปสู่การเปลี่ยนแปลงเชิงโครงสร้างทั่วไปในโมเลกุลไบโอโพลีเมอร์
myoglobin- ประกอบด้วยสายโซ่โพลีเปปไทด์หนึ่งสาย รวมถึงกรดอะมิโน 153 เรซิดิว และกลุ่มพอร์ไฟรินของเหล็ก (ฮีม) หนึ่งกลุ่มต่อโมเลกุล Myoglobin หมายถึง hemoproteins ที่สามารถจับออกซิเจนย้อนกลับได้ ในเซลล์กล้ามเนื้อโครงร่างนั้นมีหน้าที่จองออกซิเจนตลอดจนเพิ่มอัตราการแพร่ผ่านเซลล์ ทางสายวิวัฒนาการ myoglobin เป็นสารตั้งต้นของเฮโมโกลบิน โมเลกุลนี้ไม่มีพันธะไดซัลไฟด์และมีลักษณะเป็นเฮลิซิตี้ 77% heme ที่ทำหน้าที่จับกับออกซิเจนนั้นอยู่ใน "กระเป๋าที่ไม่ชอบน้ำ" ซึ่งเกิดจากกรดอะมิโนพิเศษที่มีจุดประสงค์เพื่อการนี้ Heme เป็นโปรโตพอร์ไฟรินมาโครไซเคิลที่มีไอออนเหล็กที่มีพันธะประสานกันซึ่งอยู่ตรงกลางของโมเลกุล การตรึงเชิงพื้นที่ของ heme ทำให้สามารถจับโมเลกุลออกซิเจนเป็นแกนด์ที่หกได้
เฮโมโกลบิน- โปรตีนในเลือด "ทางเดินหายใจ" มันขนส่งออกซิเจนผ่านระบบไหลเวียนโลหิตของปอดไปยังอวัยวะอื่นและศูนย์การบริโภค โมเลกุลของเฮโมโกลบินประกอบด้วยสายพอลิเปปไทด์ที่เหมือนกันเป็นคู่สี่สาย ซึ่งแต่ละสายมีเฮม สายโพลีเปปไทด์ของเฮโมโกลบินเรียกว่า a และ b , และโครงสร้างสมมาตรของโมเลกุลเขียนเป็น 2 b 2 . การก่อตัวของโครงสร้างควอเทอร์นารีดำเนินการโดยปฏิกิริยาที่ไม่ชอบน้ำระหว่างแต่ละสายโซ่โพลีเปปไทด์ เมื่อเติมออกซิเจนลงใน heme จะเกิด oxyhemoglobin โครงสร้างควอเทอร์นารีซึ่งแตกต่างจากรูปแบบที่ไม่มีออกซิเจนเพียงเล็กน้อยเท่านั้น
การเติมออกซิเจนทำให้เกิดการเปลี่ยนแปลงเชิงโครงสร้างในโมเลกุล Hb การจับกันของออกซิเจนกับการถ่ายโอนไอออน Fe 2+ ไปยังสถานะการหมุนต่ำนั้นมาพร้อมกับการกระจัดของเหล็กพร้อมกันในระนาบของกลุ่ม heme มีการแตกร้าวทีละน้อยของสะพานเกลือระหว่างหน่วยย่อย ระยะห่างระหว่าง hemes ของ a-subunits เพิ่มขึ้น และระหว่าง hemes ของ b-subunits จะลดลง โดยทั่วไป การเติมออกซิเจนจะเปลี่ยนแต่ละหน่วยย่อยจากรูปแบบดีออกซีและออกซี การแตกของสะพานเกลือสี่อันจากทั้งหมดหกอันในระหว่างการให้ออกซิเจนของหน่วยย่อย a-sub สองอันแรกมีส่วนทำให้เกิดการแตกของสะพานอีกสองอันที่เหลือ ดังนั้นจึงอำนวยความสะดวกในการยึดโมเลกุลออกซิเจนต่อไปนี้กับหน่วยย่อยที่เหลือ เพิ่มความใกล้ชิดกับออกซิเจนโดย หลายร้อยครั้ง นี่คือลักษณะสหกรณ์ของการภาคยานุวัติ
5(1). โครงสร้างปฐมภูมิและทุติยภูมิของ DNA กฎของชาร์กาฟฟ์ หลักการเสริม ประเภทของพันธะในโมเลกุลดีเอ็นเอ บทบาททางชีวภาพของดีเอ็นเอ โรคระดับโมเลกุลเป็นผลมาจากการกลายพันธุ์ของยีน
โครงสร้างเบื้องต้นของ DNA -ลำดับการสลับของดีออกซีไรโบนิวคลีโอไซด์โมโนฟอสเฟต (dNMP) ในสายพอลินิวคลีโอไทด์
หมู่ฟอสเฟตแต่ละหมู่ในสายโซ่พอลินิวคลีโอไทด์ ยกเว้นฟอสฟอรัสตกค้างที่ปลายโมเลกุล 5 "- มีส่วนร่วมในการก่อตัวของพันธะเอสเทอร์สองพันธะที่เกี่ยวข้องกับอะตอมคาร์บอน 3"- และ 5" ของดีออกซีไรโบสที่อยู่ใกล้เคียงกันสองตัว ดังนั้น พันธะระหว่างโมโนเมอร์แสดงด้วย 3", 5"- ฟอสโฟไดสเตอร์
ขั้วนิวคลีโอไทด์ของ DNA นั้นโดดเด่นด้วยโครงสร้าง: ที่ปลาย 5 "มีกลุ่มฟอสเฟตและที่ปลายสาย 3" มีกลุ่ม OH อิสระ ปลายเหล่านี้เรียกว่าปลาย 5 "และ 3" ลำดับเชิงเส้นของดีออกซีไรโบนิวคลีโอไทด์ในสายโซ่พอลิเมอร์ดีเอ็นเอมักจะย่อโดยใช้รหัสตัวอักษรเดียว ตัวอย่างเช่น -A-G-C-T-T-A-C-A- จากปลาย 5 "- ถึง 3"
โมโนเมอร์กรดนิวคลีอิกแต่ละตัวมีกรดฟอสฟอริกตกค้าง ที่ pH 7 กลุ่มฟอสเฟตจะแตกตัวเป็นไอออนอย่างเต็มที่ ดังนั้น ในร่างกายกรดนิวคลีอิกมีอยู่ในรูปของโพลิแอนไอออน (มีประจุลบหลายตัว) สารตกค้างของเพนโทสยังแสดงคุณสมบัติชอบน้ำ เบสไนโตรเจนเกือบจะไม่ละลายในน้ำ แต่อะตอมของวงแหวน purine และ pyrimidine บางส่วนสามารถเกิดขึ้นได้ พันธะไฮโดรเจน
โครงสร้างรองดีเอ็นเอ.ในปี 1953 J. Watson และ F. Crick ได้เสนอแบบจำลองโครงสร้างเชิงพื้นที่ของ DNA ตามแบบจำลองนี้ โมเลกุลดีเอ็นเอมีรูปร่างเป็นเกลียวที่เกิดจากสายโซ่พอลินิวคลีโอไทด์สองสายที่บิดเบี้ยวสัมพันธ์กันและรอบแกนร่วม เกลียวคู่ มือขวา,โซ่โพลีนิวคลีโอไทด์อยู่ในนั้น ขนานกัน(รูปที่ 4-6) เช่น ถ้าหนึ่งในนั้นอยู่ในทิศทาง 3"→5" อันที่สองจะถูกวางในทิศทาง 5"→3" ดังนั้นในแต่ละด้าน
ข้าว. 4-6. เกลียวคู่ของดีเอ็นเอโมเลกุลดีเอ็นเอประกอบด้วยสายคู่ขนานสองเส้นที่มีลำดับนิวคลีโอไทด์เสริม โซ่จะบิดสัมพันธ์กันในเกลียวขวาเพื่อให้มีเบสคู่ละประมาณ 10 คู่ต่อเทิร์น
โมเลกุล DNA อยู่ที่ปลายสายหนึ่งขนาด 5 นิ้ว และปลายอีกเส้น 3 นิ้ว
เบสของสายโซ่ DNA ทั้งหมดอยู่ภายในเกลียวคู่ และกระดูกสันหลังของเพนโทสฟอสเฟตอยู่ด้านนอก สายพอลินิวคลีโอไทด์ถูกยึดสัมพันธ์กันเนื่องจากพันธะไฮโดรเจนระหว่างเบสไนโตรเจนที่เป็นพิวรีนและไพริมิดีน A และ T (สองพันธะ) และระหว่าง G และ C (สามพันธะ) (รูปที่ 4-7) ด้วยการรวมกันนี้แต่ละ
ข้าว. 4-7. คู่เบสพิวรีน-ไพริมิดีนใน DNA
ทั้งคู่ประกอบด้วยวงแหวนสามวง ดังนั้นขนาดรวมของคู่เบสเหล่านี้จึงเท่ากันตลอดความยาวทั้งหมดของโมเลกุล พันธะไฮโดรเจนกับเบสอื่นๆ รวมกันเป็นคู่เป็นไปได้ แต่จะอ่อนกว่ามาก ลำดับนิวคลีโอไทด์ของสายเดียวประกอบขึ้นอย่างสมบูรณ์กับลำดับนิวคลีโอไทด์ของสายที่สอง ดังนั้นตามกฎของ Chargaff (Erwin Chargaff ในปี 1951 ได้กำหนดรูปแบบในอัตราส่วนของฐาน purine และ pyrimidine ในโมเลกุล DNA) จำนวนฐานของ purine (A + G) จะเท่ากับจำนวนฐานของ pyrimidine (T + C ).
ฐานเสริมจะซ้อนกันอยู่ที่แกนกลางของเกลียว ระหว่างฐานของโมเลกุลที่มีเกลียวคู่ในกอง ปฏิกิริยาที่ไม่ชอบน้ำทำให้เกลียวคู่มีเสถียรภาพ
โครงสร้างดังกล่าวไม่รวมการสัมผัสของไนโตรเจนที่ตกค้างกับน้ำ แต่ปึกฐานไม่สามารถตั้งในแนวตั้งได้อย่างสมบูรณ์ คู่เบสจะออฟเซ็ตกันเล็กน้อย ในโครงสร้างที่ขึ้นรูป ร่องสองร่องมีความโดดเด่น - ร่องขนาดใหญ่กว้าง 2.2 นาโนเมตรและร่องเล็กกว้าง 1.2 นาโนเมตร เบสไนโตรเจนในบริเวณร่องใหญ่และร่องเล็กมีปฏิสัมพันธ์กับโปรตีนเฉพาะที่เกี่ยวข้องกับการจัดโครงสร้างโครมาติน
กฎของ Chargaff- ระบบของกฎเกณฑ์เชิงประจักษ์ที่อธิบายความสัมพันธ์เชิงปริมาณระหว่างเบสไนโตรเจนชนิดต่างๆ ใน DNA พวกมันถูกคิดค้นขึ้นจากผลงานของกลุ่มนักชีวเคมี Erwin Chargaff ในปี 1949-1951
ก่อนการทำงานของกลุ่ม Chargaff ทฤษฎีที่เรียกว่า "tetranucleotide" ครอบงำตามที่ DNA ประกอบด้วยบล็อกซ้ำของสี่ฐานไนโตรเจนที่แตกต่างกัน (adenine, thymine, guanine และ cytosine) Chargaff และเพื่อนร่วมงานสามารถแยก DNA nucleotides โดยใช้ paper chromatography และหาอัตราส่วนเชิงปริมาณที่แน่นอนของ nucleotides ประเภทต่างๆ พวกมันแตกต่างอย่างมีนัยสำคัญจากฐานที่เท่ากันซึ่งคาดว่าจะได้ถ้าทั้งสี่ฐานมีสัดส่วนเท่ากัน ความสัมพันธ์ที่ระบุโดย Chargaff สำหรับ adenine (A), thymine (T), guanine (G) และ cytosine (C) มีดังนี้:
1. ปริมาณอะดีนีนเท่ากับปริมาณไทมีน และกวานีนเท่ากับไซโตซีน: A=T, G=C
2. จำนวนพิวรีนเท่ากับจำนวนพิริมิดีน: A + G = T + C
3. จำนวนเบสที่มีหมู่อะมิโนในตำแหน่ง 6 เท่ากับจำนวนเบสที่มีหมู่คีโตในตำแหน่ง 6: A+C=G+T
อย่างไรก็ตาม อัตราส่วน (A + T): (G + C) อาจแตกต่างกันใน DNA ประเภทต่างๆ. ในบางคู่ AT จะมีอำนาจเหนือกว่า ในบางคู่ - HC
กฎของ Chargaff พร้อมด้วยข้อมูลการเลี้ยวเบนของรังสีเอกซ์ มีบทบาทสำคัญในการถอดรหัสโครงสร้างของ DNA โดย J. Watson และ Francis Crick
เสริม(ใน เคมี, อณูชีววิทยา และ พันธุศาสตร์) - ความสอดคล้องกันของโมเลกุล ไบโอโพลีเมอร์หรือชิ้นส่วนของพวกมัน ซึ่งทำให้แน่ใจถึงการก่อตัวของพันธะระหว่างชิ้นส่วนเสริมเชิงพื้นที่ (เสริม) ของโมเลกุลหรือชิ้นส่วนโครงสร้างของพวกมันอันเนื่องมาจาก ปฏิกิริยาระหว่างโมเลกุล(การก่อตัวของพันธะไฮโดรเจน ปฏิกิริยาที่ไม่ชอบน้ำ ปฏิกิริยาทางไฟฟ้าสถิตของหมู่ฟังก์ชันที่มีประจุ ฯลฯ )
อันตรกิริยาของชิ้นส่วนเสริมหรือไบโอโพลีเมอร์ไม่ได้มาพร้อมกับการก่อตัวของโควาเลนต์ พันธะเคมีระหว่างชิ้นส่วนประกอบ แต่เนื่องจากการติดต่อกันเชิงพื้นที่ของชิ้นส่วนเสริมจึงนำไปสู่การก่อตัวของพันธะที่ค่อนข้างอ่อนแอจำนวนมาก (ไฮโดรเจนและ van der Waals) ด้วยพลังงานทั้งหมดขนาดใหญ่เพียงพอซึ่งนำไปสู่การก่อตัวของโมเลกุลเชิงซ้อนที่เสถียร
ในเวลาเดียวกัน ควรสังเกตว่ากลไกของกิจกรรมการเร่งปฏิกิริยาของเอนไซม์นั้นพิจารณาจากความสมบูรณ์ของเอนไซม์และสถานะการเปลี่ยนแปลงหรือผลิตภัณฑ์ขั้นกลางของปฏิกิริยาที่เร่งปฏิกิริยา - และในกรณีนี้ การก่อตัวของพันธะเคมีแบบย้อนกลับได้ เกิดขึ้น.
กรดนิวคลีอิกเสริม
เมื่อไหร่ กรดนิวคลีอิก- ทั้งเบสโอลิโก- และพอลินิวคลีโอไทด์ไนโตรเจน นิวคลีโอไทด์ได้เพราะการศึกษา พันธะไฮโดรเจนฟอร์มคู่เชิงซ้อน อะดีนีน-ไทมีน(หรือ uracilใน RNA) และ guanine-ไซโตซีนเมื่อโซ่โต้ตอบกัน กรดนิวคลีอิก. ปฏิสัมพันธ์นี้มีบทบาทสำคัญในจำนวนของ กระบวนการพื้นฐานการจัดเก็บและการส่งข้อมูลทางพันธุกรรม: การจำลองดีเอ็นเอซึ่งรับรองการถ่ายโอนข้อมูลทางพันธุกรรมระหว่างการแบ่งเซลล์ การถอดความ DNA เป็น RNA ในระหว่างการสังเคราะห์ โปรตีนเข้ารหัสโดย DNA ยีน, การจัดเก็บข้อมูลทางพันธุกรรมใน DNA แบบสองสายและกระบวนการซ่อมแซม DNA เมื่อเกิดความเสียหาย
หลักการของการเติมเต็มใช้ในการสังเคราะห์ดีเอ็นเอ นี่คือการติดต่ออย่างเข้มงวดของสารประกอบของเบสไนโตรเจนที่เชื่อมต่อด้วยพันธะไฮโดรเจนซึ่ง: A-T ( อะดีนีนเชื่อมต่อกับ thymin) จี-ซี ( Guanineเชื่อมต่อกับ ไซโตซีน)
ตัวเร่งปฏิกิริยาของเอนไซม์
การจับคู่ระหว่างเอนไซม์กับสารตั้งต้นเป็นปัจจัยสำคัญในกลไกการทำงานของเอนไซม์ และในทางตรงกันข้ามกับสถานการณ์ที่อธิบายไว้ข้างต้นด้วยการก่อตัวของสารเชิงซ้อนที่ไม่ผูกมัดทางเคมี สามารถนำไปสู่การเริ่มต้น ปฏิกิริยาเคมี- ในกรณีที่มีการเชื่อมต่อ เอนไซม์ด้วยสารตั้งต้นความสมบูรณ์ค่อนข้างต่ำอย่างไรก็ตามด้วยสถานะปฏิกิริยาการเปลี่ยนแปลงของสารตั้งต้นสูงสถานะนี้จะเสถียรซึ่งนำไปสู่ผลกระทบของกิจกรรมเร่งปฏิกิริยาของเอนไซม์: การรักษาเสถียรภาพของสถานะการเปลี่ยนแปลงนั้นเทียบเท่ากับ ลดลงใน พลังงานกระตุ้นและอัตราการเกิดปฏิกิริยาเพิ่มขึ้นอย่างรวดเร็ว
Ppt%5C34928-slozhnye_belki_ch1_1.jpg" alt="(!LANG:>ศูนย์กลางการทำงานของโปรตีนและปฏิกิริยากับลิแกนด์ ในกระบวนการสร้างโครงสร้างตติยภูมิ"> Активный центр белка и его взаимодействие с лигандом. В процессе формирования третичной структуры на поверхности функционально !} โปรตีนที่ใช้งานโดยปกติแล้วจะอยู่ในช่อง ไซต์จะถูกสร้างขึ้นจากอนุมูลของกรดอะมิโนที่อยู่ห่างไกลกันในโครงสร้างหลัก ไซต์นี้ซึ่งมีโครงสร้างเฉพาะสำหรับโปรตีนที่กำหนดและสามารถโต้ตอบอย่างจำเพาะกับโมเลกุลบางตัวหรือกลุ่มของโมเลกุลที่คล้ายคลึงกัน เรียกว่าไซต์การจับโปรตีนที่มีลิแกนด์หรือไซต์ที่ทำงานอยู่ แกนด์เป็นโมเลกุลที่ทำปฏิกิริยากับโปรตีน
Ppt%5C34928-slozhnye_belki_ch1_2.jpg" alt="(!LANG:>ลิแกนด์สามารถเป็นสารที่มีน้ำหนักโมเลกุลต่ำหรือมีน้ำหนักโมเลกุลสูง (macromolecule) ได้แก่"> Лигандом может быть как низкомолекулярное, так и высокомолекулярное (макромолекула) вещество, в том числе и другой белок. Лигандами являются субстраты ферментов, кофакторы, ингибиторы и активаторы ферментов, протомеры в олигомерном белке и т.д.!}
Ppt%5C34928-slozhnye_belki_ch1_3.jpg" alt="(!LANG:>ความจำเพาะสูงของปฏิกิริยาระหว่างโปรตีนกับลิแกนด์นั้นมั่นใจได้ด้วยการเสริมโครงสร้างของศูนย์กลางแอคทีฟที่มีโครงสร้างของลิแกนด์">!}
Ppt%5C34928-slozhnye_belki_ch1_4.jpg" alt="(!LANG:>ส่วนประกอบเสริมคือความสอดคล้องเชิงพื้นที่และทางเคมีของพื้นผิวที่มีปฏิสัมพันธ์กัน ศูนย์ที่ใช้งานอยู่ไม่ควรเพียงเท่านั้น"> Комплементарность - это пространственное и химическое соответствие взаимодействующих поверхностей. Активный центр должен не только пространственно соответствовать входящему в него лиганду, но и между функциональными группами радикалов, входящих в активный центр, и лигандом должны образоваться связи чаще всего нековалентные (ионные, водородные, а также гидрофобные взаимодействия), которые удерживают лиганд в активном центре.!}
Ppt%5C34928-slozhnye_belki_ch1_5.jpg" alt="(!LANG:>ปฏิสัมพันธ์ระหว่างโปรตีนกับลิแกนด์เสริม">!}
Ppt%5C34928-slozhnye_belki_ch1_6.jpg" alt=">">
Ppt%5C34928-slozhnye_belki_ch1_7.jpg" alt=">">
Ppt%5C34928-slozhnye_belki_ch1_8.jpg" alt="(!LANG:>การจำแนกโปรตีน 1. โปรตีนอย่างง่ายประกอบด้วยกรดอะมิโนเท่านั้น 2. โปรตีนเชิงซ้อน (โฮโลโปรตีน)"> КЛАССИФИКАЦИЯ БЕЛКОВ 1. Простые белки состоят только из аминокислот. 2. Сложные белки (холопротеины) содержат белковую часть (апопротеин) и небелковую (простетическую) группу.!}
Ppt%5C34928-slozhnye_belki_ch1_9.jpg" alt="(!LANG:>สารอินทรีย์ต่างๆ (ไขมัน คาร์โบไฮเดรต) และอนินทรีย์ (โลหะ) สามารถทำหน้าที่เป็นกลุ่มเทียมได้"> В качестве простетической группы могут выступать различные органические (липиды, углеводы) и неорганические (металлы) вещества. Связь между простетической группой и апопротеином может быть как ковалентная, так и нековалентная. Простетическую группу порой можно рассматривать в качестве лиганда. Наличие небелковой части обеспечивает выполнение белком его функции. При утрате простетической группы холопротеин теряет свою активность.!}
Ppt%5C34928-slozhnye_belki_ch1_10.jpg" alt="(!LANG:>โปรตีนเชิงซ้อน - โครโมโปรตีน - นิวคลีโอโปรตีน - ไลโปโปรตีน - ฟอสโฟโปรตีน - ไกลโคโปรตีน - เมทัลโลโปรตีน">!}
Ppt%5C34928-slozhnye_belki_ch1_11.jpg" alt="(!LANG:>Metalloproteins ประกอบด้วย holoenzymes ที่มีไอออนของโลหะที่ไม่มีการประสานกันของ heme Metalloproteins ประกอบด้วยโปรตีน"> Металлопротеинам можно отнести холоферменты, содержащие негемовые координационно связанные ионы металлов. Среди металлопротеинов есть белки, выполняющие депонирующие и транспортные функции (например, железосодержащие ферритин и трансферрин) и ферменты (например, цинксодержащая карбоангидраза и различные супероксиддисмутазы, содержащие в качестве активных центров ионы меди, марганца, железа и других металлов). Но и хромопротеины, содержащие ионы металлов, также можно отнести к металлопротеинам.!}
Ppt%5C34928-slozhnye_belki_ch1_12.jpg" alt="(!LANG:>Metalloproteins มักเป็นเอนไซม์ ในกรณีนี้ ไอออนของโลหะ: - มีส่วนร่วมในการปฐมนิเทศซับสเตรต""> Металлопротеины часто являются ферментами. Ионы металлов в этом случае: - участвуют в ориентации субстрата в активном центре фермента, входят в состав активного центра фермента и участвуют в катализе, являясь, например, акцепторами электронов на определенной стадии ферментативной реакции. Часто ион металла в составе фермента называют кофактором.!}
Ppt%5C34928-slozhnye_belki_ch1_13.jpg" alt="(!LANG:>Enzymatic metalloproteins ประกอบด้วยโปรตีนที่ประกอบด้วย: - copper - cytochrome oxidase ในคอมเพล็กซ์"> К ферментативным металлопротеинам относятся белки, содержащие например: - медь – цитохромоксидаза, в комплексе с другими ферментами дыхательной цепи митохондрий участвует в синтезе АТФ, - железо – ферритин, депонирующий железо в клетке, трансферрин, переносящий железо в крови, каталаза, обезвреживающая перекись водорода, - цинк – алкогольдегидрогеназа, обеспечивающая метаболизм этанола и других спиртов, лактатдегидрогеназа, участвующая в метаболизме молочной кислоты, - карбоангидраза, образующая угольную кислоту из CO2 и H2O, - щелочная фосфатаза, гидролизующая фосфорные эфиры различных соединений, - α2-макроглобулин, антипротеазный белок крови. - селен – тиреопероксидаза, участвующая в синтезе гормонов щитовидной железы, антиоксидантный фермент глутатионпероксидаза, - кальций – α-амилаза слюны и панкреатического сока, гидролизующая крахмал.!}
Ppt%5C34928-slozhnye_belki_ch1_14.jpg" alt="(!LANG:>เฟอร์ริติน">!}
Ppt%5C34928-slozhnye_belki_ch1_15.jpg" alt="(!LANG:>ฟอสโฟโปรตีนเป็นโปรตีนที่มีหมู่ฟอสเฟต มันจับกับสายเปปไทด์"> Фосфопротеины – это белки, в которых присутствует фосфатная группа. Она связывается с пептидной цепью через остатки тирозина, серина и треонина, т.е. тех аминокислот, которые содержат ОН-группу. Способ присоединения фосфата к белку на примере серина и тирозина!}
Ppt%5C34928-slozhnye_belki_ch1_16.jpg" alt="(!LANG:>กรดฟอสฟอริกสามารถทำหน้าที่: - บทบาทโครงสร้าง ให้ประจุ ความสามารถในการละลาย และคุณสมบัติที่เปลี่ยนแปลง"> Фосфорная кислота может выполнять: - Структурную роль, придавая заряд, растворимость и изменяя свойства белка, например, в казеине молока, яичном альбумине. Наличие остатков фосфорной кислоты способствует связыванию кальция, что необходимо для формирования, например, костной ткани. - Функциональную роль. В клетке присутствует много белков, которые связаны с фосфатом не постоянно, а в зависимости от активности метаболизма. Белок может многократно переходить в фосфорилированную или в дефосфорилированную форму, что играет регулирующую роль в его работе.!}
Ppt%5C34928-slozhnye_belki_ch1_17.jpg" alt="(!LANG:>ฟอสฟอรีเลชั่นเป็นกระบวนการถ่ายโอนกรดฟอสฟอริกตกค้างจากสารฟอสโฟรีเลตจากผู้บริจาคไปยังซับสเตรต โดยปกติ"> Фосфорилирование - процесс переноса остатка фосфорной кислоты от фосфорилирующего агента-донора к субстрату, как правило, катализируемый ферментами (киназами) и ведущий к образованию эфиров фосфорной кислоты. Дефосфорилирование (утрату остатка фосфорной кислоты) катализируют фосфатазы. АТФ + R-OH → АДФ + R-OPO3H2 R-OPO3H2 + Н2О → R-OH + Н3РО4!}
Ppt%5C34928-slozhnye_belki_ch1_18.jpg" alt="(!LANG:> ตัวอย่าง: 1) เอ็นไซม์ ไกลโคเจน ซินเทสและไกลโคเจน ฟอสโฟรีเลส 2) ฮิสโตนในสถานะฟอสโฟรีเลตมีแรงยึดเหนี่ยวน้อยกว่า"> Примеры: 1) ферменты гликогенсинтаза и гликогенфосфорилаза 2) гистоны в фосфорилированном состоянии менее прочно связываются с ДНК и активность генома возрастает. Изменение конформации белка в фосфорилированном и дефосфорилированном состоянии!}
Ppt%5C34928-slozhnye_belki_ch1_19.jpg" alt="(!LANG:>ไลโปโปรตีนประกอบด้วยไขมันที่ไม่มีพันธะโควาเลนต์เป็นส่วนเทียม โดยเฉพาะไขมัน""> Липопротеины содержат в качестве простетической части нековалентно связанные липиды. Липиды, в частности жиры, холестерол и его эфиры не растворяются в водных фазах организма, поэтому транспорт их кровью и лимфой осуществляется в виде комплексов с белками и фосфолипидами, которые называются липопротеинами.!}
Ppt%5C34928-slozhnye_belki_ch1_20.jpg" alt="(!LANG:>ไลโปโปรตีนทั้งหมดมีโครงสร้างคล้ายกัน: แกนประกอบด้วยโมเลกุลที่ไม่ชอบน้ำ: ไตรเอซิลกลีเซอรอล, เอสเทอร์ของคอเลสเตอรอล และ"> Все липопротеины имеют сходное строение: ядро состоит из гидрофобных молекул: триацилглицеролов, эфиров холестерола, а на поверхности находится монослой фосфолипидов, полярные группы которых обращены к воде, а гидрофобные погружены в гидрофобное ядро липопротеина. Кроме фосфолипидов, на поверхности находятся белки – аполипопротеины (апобелками). Их выделяют несколько видов: А, В, С, D. В каждом типе липопротеинов преобладают соответствующие ему апобелки. Аполипопротеины выполняют различные функции. Интегральные аполипопротеины являются структурными компонентами. Периферические аполипопротеины в плазме крови могут передаваться от одного типа липопротеинов к другим, определяя их дальнейшие превращения.!}
Ppt%5C34928-slozhnye_belki_ch1_21.jpg" alt="(!LANG:>แผนผังโครงสร้างไลโปโปรตีน โครงสร้างไลโปโปรตีน">!}
Ppt%5C34928-slozhnye_belki_ch1_22.jpg" alt="(!LANG:>โครงสร้างของไลโปโปรตีนในพลาสมาในเลือด">!}
Ppt%5C34928-slozhnye_belki_ch1_23.jpg" alt="(!LANG:> ไลโปโปรตีนมีสี่กลุ่มหลัก: ไลโปโปรตีนความหนาแน่นสูง (HDL), ไลโปโปรตีนความหนาแน่นต่ำ (LDL),"> Выделяют четыре основных класса липопротеинов: -липопротеины высокой плотности (ЛПВП), -липопротеины низкой плотности (ЛПНП), -липопротеины очень низкой плотности (ЛПОНП), -хиломикроны (ХМ). Каждый из типов ЛП образуется в разных тканях и транспортирует определённые липиды. Концентрация и соотношение в крови тех или иных липопротеинов играют ведущую роль в возникновении такой распространенной сосудистой патологии как атеросклероз. ЛПВП являются антиатерогенными, ЛПНП и ЛПОНП – атерогенными.!}
Ppt%5C34928-slozhnye_belki_ch1_24.jpg" alt=">">
Ppt%5C34928-slozhnye_belki_ch1_25.jpg" alt="(!LANG:>Glycoproteins หรือ glycoconjugates เป็นโปรตีนที่มีส่วนประกอบคาร์โบไฮเดรตที่เกาะติดกับกระดูกสันหลังของโพลีเปปไทด์อย่างโควาเลนต์"> Гликопротеины или, гликоконъюгаты – это белки, содержащие углеводный компонент, ковалентно присоединенный к полипептидной основе. Содержание углеводов в них варьирует от 1% до 98% по массе. Два подкласса белков, содержащих углеводы: ■ протеогликаны ■ гликопротеины!}
Description="">
Ppt%5C34928-slozhnye_belki_ch1_27.jpg" alt="(!LANG:> Glycoproteins มีคาร์โบไฮเดรดที่มีปริมาณคาร์โบไฮเดรตต่ำซึ่งติดอยู่: - โดยพันธะ N-glycosidic กับกลุ่ม NH2 ของบางส่วน"> Для гликопротеинов характерно невысокое содержание углеводов, которые присоединены: - N-гликозидной связью к NН2-группе какого-нибудь аминокислотного остатка, например, аспарагина; - О-гликозидной связью к гидроксильной группе остатка серина, треонина,тирозина, гидроксилизина.!}
Ppt%5C34928-slozhnye_belki_ch1_28.jpg" alt="(!LANG:>การก่อตัวของพันธะ O- และ N-glycosidic ในไกลโคโปรตีน 1 - พันธะ N-glycosidic ระหว่างกลุ่มเอไมด์"> Образование О- и N-гликозидных связей в гликопротеинах. 1 - N-гликозидная связь между амидной группой аспарагина и ОН-группой моносахарида; 2 - О-гликозидная связь между ОН-группой серина и ОН-группой моносахарида.!}
Ppt%5C34928-slozhnye_belki_ch1_29.jpg" alt="(!LANG:>วิธีเชื่อมโยงคาร์โบไฮเดรตเข้ากับโปรตีน">!}
Ppt%5C34928-slozhnye_belki_ch1_30.jpg" alt="(!LANG:>หน้าที่ของไกลโคโปรตีนคือ 1. โครงสร้าง - ผนังเซลล์แบคทีเรีย เมทริกซ์กระดูก เช่น คอลลาเจน อีลาสติน""> Функцией гликопротеинов являются: 1. Структурная – клеточная стенка бактерий, костный матрикс, например, коллаген, эластин. 2. Защитная – например, антитела, интерферон, факторы свертывания крови (протромбин, фибриноген). 3. Рецепторная – присоединение эффектора приводит к изменению конформации белка-рецептора, что вызывает внутриклеточный ответ. 4. Гормональная – гонадотропный, адренокортикотропный и тиреотропный гормоны. 5. Ферментативная – холинэстераза, нуклеаза. 6. Транспортная – перенос веществ в крови и через мембраны, например, трансферрин, транскортин, альбумин, Na+,К+-АТФаза.!}
Ppt%5C34928-slozhnye_belki_ch1_31.jpg" alt="(!LANG:>โครงร่างของโครงสร้างโปรตีนตัวรับ">!}
Ppt%5C34928-slozhnye_belki_ch1_32.jpg" alt="(!LANG:>โครโมโปรตีนเป็นชื่อเรียกรวมของโปรตีนเชิงซ้อนที่มีกลุ่มเทียมสีที่มีลักษณะทางเคมีต่างๆ"> Хромопротеины - собирательное название сложных белков с окрашенными простетическими группами различной химической природы. гемопротеины (содержат гем), ретинальпротеины (содержат витамин А), флавопротеины (содержат витамин В2), кобамидпротеины (содержат витамин В12).!}
Ppt%5C34928-slozhnye_belki_ch1_33.jpg" alt="(!LANG:>ฟลาโวโปรตีนเป็นเอนไซม์ของปฏิกิริยารีดอกซ์ พวกมันมีอนุพันธ์ของวิตามิน B2 ฟลาวิน โมโนนิวคลีโอไทด์ (FMN) และฟลาวิน อะดีนีน ไดนิวคลีโอไทด์""> Флавопротеины - это ферменты окислительно-восстановительных реакций. Содержат производные витамина В2 флавинмононуклеотид (ФМН) и флавинадениндинуклеотид (ФАД). Связываются данные простетические группы ковалентно и придают желтое окрашивание. Эти простетические группы являются производными изоаллоксазина.!}
Ppt%5C34928-slozhnye_belki_ch1_34.jpg" alt="(!LANG:>Isoalloxazine เป็นสารประกอบเฮเทอโรไซคลิก ซึ่งเป็นอนุพันธ์ของ pteridine โมเลกุล isoalloxazine ประกอบด้วยวงแหวนอะโรมาติกสามวง -"> Изоаллоксазин - гетероциклическое соединения, производное птеридина. Молекула изоаллоксазина состоит из трех ароматических колец - бензольного, пиримидинового, пиразинового.!}
Ppt%5C34928-slozhnye_belki_ch1_35.jpg" alt="(!LANG:>Hemoproteins เป็นโครโมโปรตีนที่มี heme ซึ่งประกอบด้วยเหล็กหรือแมกนีเซียม porphyrins ที่คล้ายคลึงกันเป็นส่วนประกอบที่ไม่ใช่โปรตีน"> Гемопротеины - гем-содержащие хромопротеины. В качестве небелкового компонента включают структурно сходные железо- или магнийпорфирины. Белковый компонент может быть разнообразным как по составу, так и по структуре. Основу структуры простетической группы большинства гемосодержащих белков составляет порфириновое кольцо, являющееся в свою очередь производным тетрапиррольного соединения – порфирина. Порфирин!}
Ppt%5C34928-slozhnye_belki_ch1_36.jpg" alt="(!LANG:>วงแหวนพอร์ไฟรินสามารถสร้างสารประกอบประสานกับไอออนโลหะต่างๆ ได้ อันเป็นผลมาจากการก่อตัวที่ซับซ้อน"> Порфириновое кольцо способно образовывать координационные соединения с различными ионами металлов. В результате комплексообразования формируются металлопорфирины: содержащие ионы железа – гемоглобины, миоглобин, цитохромы, пероксидаза, каталаза и др. (красное окрашивание), содержщие ионы магния – хлорофилл (зеленое окрашивание). Витамин В12 (кобалимин) содержит координированный ион кобальта Со2+ в порфириноподобном макроцикле – коррине, состоящем из четырех частично гидрированных пиррольных колец (розовое окрашивание).!}
Ppt%5C34928-slozhnye_belki_ch1_37.jpg" alt="(!LANG:>คลอโรฟิลล์ ข. คลอโรฟิลล์มีส่วนร่วมในการสังเคราะห์แสง">!}
Ppt%5C34928-slozhnye_belki_ch1_38.jpg" alt="(!LANG:> ไซโตโครมแตกต่างกันในองค์ประกอบของกรดอะมิโนของสายเปปไทด์ จำนวนสาย และแบ่งออกเป็นประเภท a, b,"> Цитохромы различаются аминокислотным составом пептидных цепей, числом цепей и разделяются на типы а, b, с, d. Цитохромы находятся в составе дыхательной цепи и цепи микросомального окисления. Степень окисления железа в составе цитохромов меняется в отличие от гемоглобина и миоглобина Fe2+ ↔ Fe3+!}
Ppt%5C34928-slozhnye_belki_ch1_39.jpg" alt="(!LANG:>Myoglobin (Mb)) เป็นโปรตีนที่พบในกล้ามเนื้อสีแดง หน้าที่หลักคือสร้างพลังงานสำรอง"> Миоглобин (Мв) - белок, находящийся в красных мышцах, основная функция которого - создание запасов О2, необходимых при интенсивной мышечной работе. Мв - сложный белок, содержащий белковую часть - апоМв и небелковую часть - гем. Первичная структура апоМв определяет его компактную глобулярную конформацию и структуру активного центра, к которому присоединяется небелковая часть миоглобина - гем. Кислород, поступающий из крови в мышцы, связывается с Fe2+ гема в составе миоглобина. Мв - мономерный белок, имеющий очень высокое сродство к О2, поэтому отдача кислорода миоглобином происходит только при интенсивной мышечной работе, когда парциальное давление O2 резко снижается. Формирование пространственных структур и функционирование миоглобина.!}
Ppt%5C34928-slozhnye_belki_ch1_40.jpg" alt="(!LANG:>รูปแบบของรูปแบบ MB"> Формирование конформации Мв. В красных мышцах на рибосомах в ходе трансляции идет синтез первичной структуры Мв, представленной специфической последовательностью 153 аминокислотных остатков. Вторичная структура Мв содержит восемь α-спиралей, называемых латинскими буквами от А до Н, между которыми имеются неспирализованные участки. Третичная структура Мв имеет вид компактной глобулы, в углублении которой между F и Е α-спиралями расположен активный центр.!}
Ppt%5C34928-slozhnye_belki_ch1_41.jpg" alt="(!LANG:>โครงสร้าง Myoglobin">!}
Ppt%5C34928-slozhnye_belki_ch1_42.jpg" alt="(!LANG:>คุณลักษณะของโครงสร้างและการทำงานของศูนย์แอคทีฟ Mv ศูนย์แอคทีฟ Mv ส่วนใหญ่เกิดจากอนุมูลที่ไม่ชอบน้ำ"> Особенности строения и функционирования активного центра Мв. Активный центр Мв сформирован преимущественно гидрофобными радикалами аминокислот, далеко отстоящими друг от друга в первичной структуре (например, Три39 и Фен138). К активному центру присоединяется плохо растворимые в воде лиганды - гем и О2. Гем - специфический лиганд апоМв.!}
Ppt%5C34928-slozhnye_belki_ch1_43.jpg" alt="(!LANG:>Heme ขึ้นอยู่กับวงแหวนไพร์โรลสี่วงที่เชื่อมต่อกันด้วยเมทิลบริดจ์ โดยตรงกลางคืออะตอม Fe2+"> Основу гема составляют четыре пиррольных кольца, соединенных метенильными мостиками; в центре расположен атом Fe2+, соединенный с атомами азота пиррольных колец четырьмя координационными связями. В активном центре Мв кроме гидрофобных радикалов аминокислот имеются также остатки двух аминокислот с гидрофильными радикалами - Гис Е7 (Гис64) и Гис F8 (Гис93).!}
Ppt%5C34928-slozhnye_belki_ch1_44.jpg" alt="(!LANG:>F8 ของเขาสร้างพันธะประสานงานกับ Fe2+ และแก้ไข heme อย่างแน่นหนาในไซต์ที่ทำงานอยู่"> Гис F8 образует координационную связь с Fe2+ и прочно фиксирует гем в активном центре. Гис Е7 необходим для правильной ориентации в активном центре другого лиганда - O2 при его взаимодействии с Fe+2 гема. Микроокружение гема создает условия для прочного, но обратимого связывания O2 с Fe+2 и препятствует попаданию в гидрофобный активный центр воды, что может привести к его окислению в Fе3+.!}
Ppt%5C34928-slozhnye_belki_ch1_45.jpg" alt="(!LANG:>โครงสร้าง Oligomeric ของ Hb และระเบียบความสัมพันธ์ของ Hb สำหรับ O2 โดยลิแกนด์ ฮีโมโกลบินของมนุษย์ -"> Олигомерное строение Нв и регуляция сродства Нв к О2 лигандами. Гемоглобины человека - семейство белков, так же как и миоглобин относящиеся к сложным белкам (гемопротеинам). Они имеют тетрамерное строение и содержат две α-цепи, но различаются по строению двух других полипептидных цепей (2α-, 2х-цепи). Строение второй полипептидной цепи определяет особенности функционирования этих форм Нв. Около 98% гемоглобина эритроцитов взрослого человека составляет гемоглобин А (2α-, 2β-цепи). В период внутриутробного развития функционируют два основных типа гемоглобинов: эмбриональный Нв (2α, 2ε), который обнаруживается на ранних этапах развития плода, и гемоглобин F (фетальный) - (2α, 2γ), который приходит на смену раннему гемоглобину плода на шестом месяце внутриутробного развития и только после рождения замещается на Нв А.!}
Ppt%5C34928-slozhnye_belki_ch1_46.jpg" alt="(!LANG:> Hb A เป็นโปรตีนที่เกี่ยวข้องกับ myoglobin (Mb)) ที่พบในเม็ดเลือดแดงของผู้ใหญ่ โครงสร้างของมัน"> Нв А - белок, родственный миоглобину (Мв), содержится в эритроцитах взрослого человека. Строение его отдельных протомеров аналогично таковому у миоглобина. Вторичная и третичная структуры миоглобина и протомеров гемоглобина очень сходны, несмотря на то что в первичной структуре их полипептидных цепей идентичны только 24 аминокислотных остатка (вторичная структура протомеров гемоглобина, так же как миоглобин, содержит восемь α-спиралей, обозначаемых латинскими буквами от А до Н, а третичная структура имеет вид компактной глобулы). Но в отличие от миоглобина гемоглобин имеет олигомерное строение, состоит из четырех полипептидных цепей, соединенных нековалентными связями.!}
Ppt%5C34928-slozhnye_belki_ch1_47.jpg" alt="(!LANG:>โครงสร้างโอลิโกเมอร์ของเฮโมโกลบิน">!}
Ppt%5C34928-slozhnye_belki_ch1_48.jpg" alt="(!LANG:>โปรโตเมอร์แต่ละ Hb เชื่อมต่อกับส่วนที่ไม่ใช่โปรตีน - heme และโปรโตเมอร์ที่อยู่ใกล้เคียง"> Каждый протомер Нв связан с небелковой частью - гемом и соседними протомерами. Соединение белковой части Нв с гемом аналогично таковому у миоглобина: в активном центре белка гидрофобные части гема окружены гидрофобными радикалами аминокислот за исключением Гис F8 и Гис Е7, которые расположены по обе стороны от плоскости гема и играют аналогичную роль в функционировании белка и связывании его с кислородом. Кроме того, Гис Е7 выполняет важную дополнительную роль в функционировании Нв. Свободный гем имеет в 25 000 раз более высокое сродство к СО, чем к О2. СО в небольших количествах образуется в организме и, учитывая его высокое сродство к гему, он мог бы нарушать транспорт необходимого для жизни клеток О2. Однако в составе гемоглобина сродство гема к оксиду углерода превышает сродство к О2 всего в 200 раз благодаря наличию в активном центре Гис Е7. Остаток этой аминокислоты создает !} เงื่อนไขที่เหมาะสมที่สุดสำหรับจับ heme กับ O2 และทำให้ปฏิกิริยาของ heme กับ CO อ่อนแอลง
Ppt%5C34928-slozhnye_belki_ch1_49.jpg" alt=">">
Ppt%5C34928-slozhnye_belki_ch1_50.jpg" alt="(!LANG:>วงแหวนฮีมไพร์โรลอยู่ในระนาบเดียวกัน และไอออน Fe2+ ในสถานะไม่มีออกซิเจนของ Hb"> Пиррольные кольца гема расположены в одной плоскости, а ион Fe2+ в неоксигенированом состоянии Hb выступает над плоскостью на 0,6 А. При присоединении кислорода ион железа погружается в плоскость колец гема. В результате сдвигается и участок полипептидной цепи, нарушаются слабые связи в молекуле Hb и изменяется конформация всей глобулы. Таким образом, присоединение кислорода вызывает изменение пространственной структуры молекулы миоглобина или протомеров гемоглобина.!}
Ppt%5C34928-slozhnye_belki_ch1_51.jpg" alt="(!LANG:>เฮโมโกลบินมีอยู่ได้ทั้งแบบอิสระ (ดีออกซีเฮโมโกลบิน)" และอยู่ในรูปของออกซิเจน"> Гемоглобин может существовать как в свободной (дезоксигемоглобин), так и в оксигенированной форме, присоединяя до 4 молекул кислорода. Взаимодействие с кислородом 1-го протомера вызывает изменение его конформации, а также кооперативные конформационные изменения остальных протомеров. Сродство к кислороду возрастает, и присоединение О2 к активному центру 2-го протомера происходит легче, вызывая дальнейшую конформационную перестройку всей молекулы. В результате еще сильнее изменяется структура оставшихся протомеров и их активных центров, взаимодействие с О2 еще больше облегчается. В итоге 4-я молекула кислорода присоединяется к Hb примерно в 300 раз легче, чем 1-я. Так происходит в легких при высоком парциальном давлении кислорода.!}
Ppt%5C34928-slozhnye_belki_ch1_52.jpg" alt="(!LANG:>ความร่วมมือการเปลี่ยนแปลงโครงสร้างของโมเลกุลเฮโมโกลบินเมื่อมีปฏิสัมพันธ์กับออกซิเจน">!}
Ppt%5C34928-slozhnye_belki_ch1_53.jpg" alt="(!LANG:> ในเนื้อเยื่อที่มีปริมาณออกซิเจนต่ำ ในทางกลับกัน การกำจัดโมเลกุล O2 แต่ละโมเลกุลช่วยให้เกิดการปล่อยโมเลกุลที่ตามมาได้ง่ายขึ้น"> В тканях, где содержание кислорода ниже, наоборот, отщепление каждой молекулы О2 облегчает освобождение последующих. Таким образом, взаимодействие олигомерного белка гемоглобина с лигандом (О2) в одном центре связывания приводит к изменению конформации всей молекулы и других, пространственно удаленных центров, расположенных на других субъединицах (принцип «домино»). Подобные взаимосвязанные изменения структуры белка называют кооперативными конформационными изменениями. Они характерны для всех олигомерных белков и используются для регуляции их активности.!}
Ppt%5C34928-slozhnye_belki_ch1_54.jpg" alt="(!LANG:>ปฏิกิริยาของโปรตีนทั้งสอง (Mb และ Hb)) กับออกซิเจนขึ้นอยู่กับความดันบางส่วนใน"> Взаимодействие обоих белков (Mb и Hb) с кислородом зависит от его парциального давления в тканях. Эта зависимость имеет разный характер, что связано с их особенностями структуры и функционирования. Гемоглобин имеет S-образную кривую насыщения, которая показывает, что субъединицы белка работают кооперативно, и чем больше кислорода они отдают, тем легче идет освобождение остальных молекул О2. Этот процесс зависит от изменения парциального давления кислорода в тканях. График насыщения миоглобина кислородом имеет характер простой гиперболы, т.е. насыщение Mb кислородом происходит быстро и отражает его функцию - обратимое связывание с кислородом, высвобождаемым гемоглобином, и освобождение в случае интенсивной физической нагрузки.!}
Ppt%5C34928-slozhnye_belki_ch1_55.jpg" alt="(!LANG:>เส้นโค้งความอิ่มตัวของออกซิเจนในเลือดและฮีโมโกลบิน">!}
Ppt%5C34928-slozhnye_belki_ch1_56.jpg" alt="(!LANG:>CO2 and H+ เกิดขึ้นระหว่างกระบวนการ catabolism ของสารอินทรีย์ ลดความสัมพันธ์ของเฮโมโกลบินกับ O2 ตามสัดส่วน""> CO2 и Н+, образующиеся при катаболизме органических веществ, уменьшают сродство гемоглобина к О2 пропорционально их концентрации. Энергия, необходимая для работы клеток, вырабатывается преимущественно в митохондриях при окислении органических веществ с использованием O2, доставляемого из легких гемоглобином. В результате окисления органических веществ образуются конечные продукты их распада: СО2 и Н2O, количество которых пропорционально интенсивности протекающих процессов окисления. СO2 диффузией попадает из клеток в кровь и проникает в эритроциты, где под действием фермента карбоангидразы превращается в угольную кислоту. Эта слабая кислота диссоциирует на протон и бикарбонат ион. СО2 + Н2О → Н2СО3 → Н+ + НСО3-!}
Ppt%5C34928-slozhnye_belki_ch1_57.jpg" alt="(!LANG:> ไอออน H+ สามารถเพิ่มอนุมูล His146 ในเฮโมโกลบิน β-chains ได้ เช่น ในพื้นที่ห่างไกลจาก"> Ионы Н+ способны присоединятся к радикалам Гис146 в β-цепях гемоглобина, т.е. в участках, удаленных от гема. Протонирование гемоглобина снижает его сродство к О2, способствует отщеплению О2 от оксиНв, образованию дезоксиНв и увеличивает поступление кислорода в ткани пропорционально количеству образовавшихся протонов. Увеличение количества освобожденного кислорода в зависимости от увеличения концентрации Н+ в эритроцитах называется эффектом Бора (по имени датского физиолога Христиана Бора, впервые открывшего этот эффект). В легких высокое парциальное давление кислорода способствует его связыванию с дезоксиНв, что уменьшает сродство белка к Н+. Освободившиеся протоны под действием карбоангидразы взаимодействуют с бикарбонатами с образованием СО2 и Н2О!}
Ppt%5C34928-slozhnye_belki_ch1_58.jpg" alt="(!LANG:>การพึ่งพาอาศัยกันของ Hb สำหรับ O2 ต่อความเข้มข้นของคาร์บอนไดออกไซด์และโปรตอน (ผลของบอร์): A -"> Зависимость сродства Нв к О2 от концентрации СО2 и протонов (эффект Бора): А - влияние концентрации СО2 и Н+ на высвобождение О2 из комплекса с Нв (эффект Бора); Б - оксигенирование дезоксигемоглобина в легких, образование и выделение СО2.!}
Ppt%5C34928-slozhnye_belki_ch1_59.jpg" alt="(!LANG:>CO2 ที่ได้จะเข้าสู่ช่องว่างและถูกขับออกด้วยอากาศที่หายใจออก ดังนั้น ปริมาณ"> Образовавшийся СО2 поступает в альвеолярное пространство и удаляется с выдыхаемым воздухом. Таким образом, количество высвобождаемого гемоглобином кислорода в тканях регулируется продуктами катаболизма органических веществ: чем интенсивнее распад веществ, например при !} การออกกำลังกายยิ่งความเข้มข้นของ CO2 และ H+ สูงขึ้น และเนื้อเยื่อได้รับออกซิเจนมากขึ้นเนื่องจากความสัมพันธ์ของ Hb สำหรับ O2 ลดลง
Ppt%5C34928-slozhnye_belki_ch1_60.jpg" alt="(!LANG:>การเปลี่ยนแปลงในการทำงานของโปรตีนเมื่อมีปฏิสัมพันธ์กับลิแกนด์อื่นๆ เนื่องจากการเปลี่ยนแปลงของโครงสร้างเรียกว่าอัลโลสเทอริก""> Изменение функциональной активности белка при взаимодействии с другими лигандами вследствие конформационных изменений называется аллостерической регуляцией, а соединения-регуляторы - аллостерическими лигандами или эффекторами. Способность к аллостерической регуляции характерна, как правило, для олигомерных белков, т.е. для проявления аллостерического эффекта необходимо взаимодействие протомеров. При воздействии аллостерических лигандов белки меняют свою конформацию (в том числе и активного центра) и функцию.!}
Ppt%5C34928-slozhnye_belki_ch1_61.jpg" alt="(!LANG:>Allosteric Regulation of Hb affinity for O2 by the 2,3-bis-phosphoglycerate ligand. ในเม็ดเลือดแดงจากผลิตภัณฑ์"> Аллостерическая регуляция сродства Нв к О2 лигандом - 2,3-бис-фосфоглицератом. В эритроцитах из продукта окисления глюкозы - 1,3-бисфосфоглицерата синтезируется аллостерический лиганд гемоглобина - 2,3-бисфосфоглицерат (2,3-БФГ). В нормальных условиях концентрация 2,3-БФГ высокая и сравнима с концентрацией Нв. 2,3-БФГ имеет сильный отрицательный заряд (-5).!}
Ppt%5C34928-slozhnye_belki_ch1_62.jpg" alt="(!LANG:>มีโพรงอยู่ตรงกลางของโมเลกุลเตตระเมอร์ของเฮโมโกลบิน ซึ่งเกิดจากกรดอะมิโนตกค้างของโปรโตเมอร์ทั้งสี่ตัว"> В центре тетрамерной молекулы гемоглобина находится полость. Ее образуют аминокислотные остатки всех четырех протомеров. В капиллярах тканей протонирование Нв (эффект Бора) приводит к разрыву связи между железом гема и О2. В молекуле дезоксигемоглобина по сравнению с оксигемоглобином возникают дополнительные ионные связи, соединяющие протомеры, вследствие чего размеры центральной полости по сравнению с оксигемоглобином увеличиваются. Центральная полость является местом присоединения 2,3-БФГ к гемоглобину. БФГ поступает в полость дезоксигемоглобина. 2,3-БФГ взаимодействует с гемоглобином в участке, удаленном от активных центров белка и относится к аллостерическим (регуляторным) лигандам, а центральная полость Нв является аллостерическим центром. 2,3-БФГ имеет сильный отрицательный заряд и взаимодействует с положительно заряженными группами двух β-цепей Нв. При этом его сродство к О2 снижается в 26 раз. В результате происходит высвобождение кислорода в капиллярах ткани при низком парциальном давлении О2. В легких высокое парциальное давление О2, наоборот, приводит к оксигенированию Нв и освобождению БФГ.!}
Ppt%5C34928-slozhnye_belki_ch1_63.jpg" alt="(!LANG:>ศูนย์รวม BPG ตั้งอยู่ในช่องที่มีประจุบวกระหว่าง 4 โปรโตเมอร์ของเฮโมโกลบิน ปฏิสัมพันธ์ของ BPG"> Центр связывания БФГ находится в положительно заряженной полости между 4 протомерами гемоглобина. Взаимодействие БФГ с центром связывания изменяет конформацию α- и β-протомеров НЬ и их активных центров. Сродство НЬ к молекулам О2 снижается и кислород высвобождается в ткани. В легких при высоком парциальном давлении О2 активные центры гемоглобина насыщаются за счет изменения конформации и БФГ вытесняется из аллостерического центра!}
Ppt%5C34928-slozhnye_belki_ch1_64.jpg" alt=">">
Ppt%5C34928-slozhnye_belki_ch1_65.jpg" alt="(!LANG:>ดังนั้น oligomeric proteins จึงมีคุณสมบัติใหม่เมื่อเปรียบเทียบกับโปรตีน monomeric การเกาะติดของลิแกนด์"> Таким образом, олигомерные белки обладают новыми по сравнению с мономерными белками свойствами. Присоединение лигандов на участках, пространственно удаленных друг от друга (аллостерических), способно вызывать конформационные изменения во всей белковой молекуле. Благодаря взаимодействию с регуляторными лигандами происходит изменение конформации и адаптация функции белковой молекулы к изменениям окружающей среды.!}
Ppt%5C34928-slozhnye_belki_ch1_66.jpg" alt="(!LANG:>ประมาณ 15% คาร์บอนไดออกไซด์ที่มีอยู่ในเลือดจะถูกขนส่งโดยโมเลกุลของเฮโมโกลบิน ในเนื้อเยื่อ โมเลกุลบางตัว "> ประมาณ 15% ของคาร์บอนไดออกไซด์ที่มีอยู่ในเลือดถูกลำเลียงโดยโมเลกุลของเฮโมโกลบิน ในเนื้อเยื่อ โมเลกุลของคาร์บอนไดออกไซด์บางส่วนสามารถเกาะกับโปรโตเมอร์แต่ละตัวของโมเลกุลเฮโมโกลบินได้ Hb สำหรับออกซิเจนลดลง ในทางกลับกัน เนื่องจากความดันบางส่วนของออกซิเจนสูง O2 จะจับกับ Hb และปล่อย CO2
Ppt%5C34928-slozhnye_belki_ch1_67.jpg" alt=">">
Ppt%5C34928-slozhnye_belki_ch1_68.jpg" alt="(!LANG:>ในโมเลกุล S ของฮีโมโกลบิน (นี่คือลักษณะที่เรียกว่าฮีโมโกลบินผิดปกติ)" 2 โซ่ β ถูกกลายพันธุ์ซึ่ง"> В молекуле гемоглобина S (так назван аномальный гемоглобин) мутантными оказались 2 β-цепи, в которых глутамат, высокополярная отрицательно заряженная аминокислота в положении 6 была заменена валином, содержащим гидрофобный радикал.!}
ความจำเพาะสูงของการจับโปรตีนกับลิแกนด์นั้นมาจากการเติมเต็มของโครงสร้างของแอคทีฟไซต์ของโปรตีนที่มีโครงสร้างของลิแกนด์
การเติมเต็มเป็นที่เข้าใจกันว่าเป็นการสอดคล้องกันเชิงพื้นที่และทางเคมีของโมเลกุลที่มีปฏิสัมพันธ์ ลิแกนด์ต้องสามารถเข้าได้และสอดคล้องกับโครงสร้างของไซต์แอ็คทีฟ ความบังเอิญนี้อาจไม่สมบูรณ์ แต่เนื่องจากความสามารถในการกำหนดโครงสร้างของโปรตีน ศูนย์ที่ทำงานอยู่สามารถเปลี่ยนแปลงเล็กน้อยและ "ปรับ" ให้กับแกนด์ได้ นอกจากนี้ ระหว่างกลุ่มหน้าที่ของลิแกนด์กับอนุมูลของกรดอะมิโนที่ก่อตัวเป็นศูนย์แอคทีฟ ควรมีพันธะที่ยึดลิแกนด์ไว้ที่ศูนย์กลางแอคทีฟ พันธะระหว่างลิแกนด์และศูนย์กลางแอคทีฟของโปรตีนอาจเป็นแบบไม่มีโควาเลนต์ (ไอออนิก ไฮโดรเจน ไม่ชอบน้ำ) หรือโควาเลนต์ก็ได้
1. ลักษณะของศูนย์ปฏิบัติการ
ศูนย์กลางที่ทำงานอยู่ของโปรตีนคือตำแหน่งที่ค่อนข้างแยกได้จากสภาพแวดล้อมรอบ ๆ โปรตีน ซึ่งก่อตัวขึ้นจากกากกรดอะมิโน ในบริเวณนี้ สารตกค้างแต่ละชนิดตามขนาดและกลุ่มการทำงานแต่ละชนิด ทำให้เกิด "การบรรเทา" ของศูนย์ที่ใช้งานอยู่
การรวมกรดอะมิโนดังกล่าวเข้าไว้ในสารเชิงซ้อนเชิงฟังก์ชันเดียวจะเปลี่ยนปฏิกิริยาของอนุมูลอิสระ เช่นเดียวกับเสียงที่เปลี่ยนไป เครื่องดนตรีในวงดนตรี ดังนั้น เรซิดิวของกรดอะมิโนที่ประกอบเป็นแอกทีฟไซต์จึงมักถูกเรียกว่า "กลุ่ม" ของกรดอะมิโน
คุณสมบัติเฉพาะของศูนย์แอคทีฟไม่เพียงขึ้นอยู่กับคุณสมบัติทางเคมีของกรดอะมิโนที่ก่อตัวเท่านั้น แต่ยังขึ้นกับการวางแนวร่วมกันที่แน่นอนในอวกาศด้วย ดังนั้นแม้การรบกวนเล็กน้อยในโครงสร้างทั่วไปของโปรตีนอันเป็นผลมาจากการเปลี่ยนแปลงจุดในโครงสร้างหลักหรือสภาวะแวดล้อมสามารถนำไปสู่การเปลี่ยนแปลงในคุณสมบัติทางเคมีและการทำงานของอนุมูลที่ก่อตัวเป็นศูนย์ที่ใช้งานอยู่ ขัดขวางการผูกมัดของ โปรตีนต่อลิแกนด์และหน้าที่ของมัน ในระหว่างการทำให้เสียสภาพ ศูนย์กลางของโปรตีนจะถูกทำลาย และกิจกรรมทางชีวภาพของพวกมันจะหายไป
ศูนย์กลางแอคทีฟมักถูกสร้างขึ้นในลักษณะที่การเข้าถึงน้ำไปยังกลุ่มการทำงานของอนุมูลถูกจำกัด เงื่อนไขถูกสร้างขึ้นสำหรับการผูกลิแกนด์กับอนุมูลของกรดอะมิโน
ในบางกรณี ลิแกนด์ติดอยู่กับอะตอมเพียงอะตอมเดียวที่มีปฏิกิริยาบางอย่าง เช่น การเพิ่ม O 2 เข้ากับเหล็กของไมโอโกลบินหรือเฮโมโกลบิน อย่างไรก็ตาม คุณสมบัติของอะตอมที่กำหนดในการเลือกโต้ตอบกับ O 2 นั้นพิจารณาจากคุณสมบัติของอนุมูลที่อยู่รอบอะตอมของเหล็กในองค์ประกอบของหัวข้อ Heme ยังพบได้ในโปรตีนอื่นๆ เช่น ไซโตโครม อย่างไรก็ตาม หน้าที่ของอะตอมของเหล็กในไซโตโครมนั้นแตกต่างกัน มันทำหน้าที่เป็นตัวกลางในการถ่ายโอนอิเล็กตรอนจากสารหนึ่งไปยังอีกสารหนึ่ง ในขณะที่ธาตุเหล็กจะกลายเป็นสารสองวาเลนต์หรือไตรวาเลนต์
คุณสมบัติหลักของโปรตีนที่อยู่ภายใต้หน้าที่ของพวกมันคือการคัดเลือกของการติดลิแกนด์จำเพาะกับบางส่วนของโมเลกุลโปรตีน
2. ลิแกนด์ที่หลากหลาย
ลิแกนด์อาจเป็นสารอนินทรีย์ (มักเป็นไอออนของโลหะ) และสารอินทรีย์ น้ำหนักโมเลกุลต่ำ และสารที่มีน้ำหนักโมเลกุลสูง
มีลิแกนด์ที่เปลี่ยนโครงสร้างทางเคมีเมื่อยึดติดกับศูนย์กลางของโปรตีน (การเปลี่ยนแปลงของสารตั้งต้นในศูนย์กลางการทำงานของเอนไซม์)
มีลิแกนด์ที่เกาะติดกับโปรตีนในขณะทำงานเท่านั้น (เช่น O 2 ขนส่งโดยเฮโมโกลบิน) และลิแกนด์ที่เกี่ยวข้องกับโปรตีนอย่างต่อเนื่องและมีบทบาทสำคัญในการทำงานของโปรตีน (เช่น เหล็ก ซึ่งเป็นส่วนหนึ่งของเฮโมโกลบิน)
ในกรณีเหล่านั้นเมื่อกรดอะมิโนที่ตกค้างซึ่งก่อตัวเป็นศูนย์ออกฤทธิ์ไม่สามารถรับรองการทำงานของโปรตีนนี้ โมเลกุลที่ไม่ใช่โปรตีนสามารถเกาะติดกับบางส่วนของศูนย์ออกฤทธิ์ได้ ดังนั้นในศูนย์กลางของเอนไซม์จำนวนมากจึงมีไอออนของโลหะ (โคแฟคเตอร์) หรือโมเลกุลที่ไม่ใช่โปรตีนอินทรีย์ (โคเอ็นไซม์) ส่วนที่ไม่ใช่โปรตีนซึ่งมีความเกี่ยวข้องอย่างยิ่งกับตำแหน่งแอคทีฟของโปรตีนและจำเป็นต่อการทำงานของมัน เรียกว่า "กลุ่มต่อมลูกหมาก" Myoglobin, hemoglobin และ cytochromes มีกลุ่มเทียมในศูนย์ที่ใช้งานอยู่ - heme ที่มีธาตุเหล็ก
การเชื่อมต่อของโปรโตเมอร์ในโปรตีนโอลิโกเมอร์เป็นตัวอย่างของการทำงานร่วมกันของลิแกนด์ที่มีน้ำหนักโมเลกุลสูง protomer แต่ละตัวที่เชื่อมต่อกับ protomers อื่น ๆ ทำหน้าที่เป็นแกนด์สำหรับพวกเขาเช่นเดียวกับที่เป็นอยู่
บางครั้งการเติมลิแกนด์จะเปลี่ยนโครงสร้างของโปรตีน ส่งผลให้เกิดการเกาะติดกับลิแกนด์อื่นๆ ตัวอย่างเช่น โปรตีนคาโมดูลิน หลังจากจับกับ Ca 2+ สี่ไอออนในพื้นที่เฉพาะ จะได้รับความสามารถในการโต้ตอบกับเอนไซม์บางชนิด ทำให้กิจกรรมของพวกมันเปลี่ยนไป
8. โครงสร้างควอเทอร์นารีของโปรตีน คุณสมบัติของโครงสร้างและการทำงานของโปรตีนโอลิโกเมอร์ในตัวอย่างของเฮโมโกลบิน การเปลี่ยนแปลงแบบมีส่วนร่วมในรูปแบบโปรโตเมอร์ ความเป็นไปได้ของการควบคุมการทำงานทางชีวภาพของโปรตีนโอลิโกเมอร์โดยลิแกนด์อัลโลสเตอริก
ภายใต้โครงสร้างควอเทอร์นารีหมายถึงวิธีการวางสายโซ่โพลีเปปไทด์แต่ละตัวในช่องว่างที่มีโครงสร้างหลัก รองหรือตติยรีเดียวกัน (หรือต่างกัน) และการก่อตัวของการก่อรูปโมเลกุลขนาดใหญ่ในลักษณะโครงสร้างและการทำงาน โปรตีนที่ใช้งานได้หลายอย่างประกอบด้วยสายพอลิเปปไทด์หลายสายที่เชื่อมต่อกันไม่ใช่ด้วยพันธะโควาเลนต์ แต่โดยพันธะที่ไม่ใช่โควาเลนต์ (คล้ายกับพันธะที่ให้ความเสถียรกับโครงสร้างตติยภูมิ) แต่ละสายโซ่โพลีเปปไทด์ เรียกว่าโปรโตเมอร์ โมโนเมอร์ หรือหน่วยย่อย ส่วนใหญ่มักไม่มีฤทธิ์ทางชีวภาพ โปรตีนได้รับความสามารถนี้ด้วยวิธีบางอย่างของการเชื่อมโยงเชิงพื้นที่ของโปรโตเมอร์ที่เป็นส่วนประกอบ เช่น มีคุณสมบัติใหม่ที่ไม่มีลักษณะเฉพาะของโปรตีนโมโนเมอร์ โมเลกุลที่เป็นผลลัพธ์มักเรียกว่าโอลิโกเมอร์ (หรือมัลติเมอร์) โปรตีนโอลิโกเมอร์มักถูกสร้างขึ้นจากโปรโตเมอร์จำนวนเท่ากัน (ตั้งแต่ 2 ถึง 4 ไม่ค่อยมีตั้งแต่ 6 ถึง 8) โดยมีน้ำหนักโมเลกุลเท่ากันหรือต่างกัน - จากหลายพันถึงหลายแสน โดยเฉพาะอย่างยิ่ง โมเลกุลของเฮโมโกลบินประกอบด้วยสาย α- ที่เหมือนกันสองสายและสาย β-โพลีเปปไทด์สองสาย กล่าวคือ เป็นเตตระเมอร์
การเปลี่ยนแปลงแบบมีส่วนร่วมในรูปแบบโปรโตเมอร์
การเปลี่ยนแปลงโครงสร้างและด้วยเหตุนี้ในคุณสมบัติเชิงหน้าที่ของโปรโตเมอร์ทั้งหมดของโปรตีนโอลิโกเมอริกเมื่อลิแกนด์ติดอยู่กับลิแกนด์เพียงตัวเดียวจึงเรียกว่าการเปลี่ยนแปลงร่วมกันในโครงสร้างของโปรโตเมอร์
ระเบียบ Allosteric . เอ็นไซม์เปลี่ยนแปลงกิจกรรมผ่านเอฟเฟกเตอร์ที่ไม่สัมพันธ์กับโควาเลนต์ การผูกมัดเกิดขึ้นในพื้นที่ ห่างไกลจากไซต์ที่ทำงานอยู่ (ตัวเร่งปฏิกิริยา) การจับนี้ทำให้เกิดการเปลี่ยนแปลงโครงสร้างในโมเลกุลโปรตีน นำไปสู่การเปลี่ยนแปลงในเรขาคณิตจำเพาะของไซต์ตัวเร่งปฏิกิริยา กิจกรรมสามารถเพิ่มขึ้น - นี่คือการเปิดใช้งานของเอนไซม์หรือลดลง - นี่คือการยับยั้ง "ข้อความ" เกี่ยวกับสิ่งที่แนบมาของตัวกระตุ้น allosteric จะถูกส่งผ่านการเปลี่ยนแปลงโครงสร้างไปยังหน่วยย่อยของตัวเร่งปฏิกิริยาซึ่งกลายเป็นส่วนประกอบเสริมของสารตั้งต้นและเอนไซม์จะถูก "เปลี่ยน" บน". เมื่อตัวกระตุ้นถูกกำจัดออกไป เอ็นไซม์จะกลับเข้าสู่สภาพที่ไม่ใช้งานอีกครั้งและ "ดับ" การควบคุม Allosteric เป็นโหมดหลักของการควบคุมเส้นทางการเผาผลาญ
โมดูลโปรตีน (โดเมน)
โดยปกติ โปรตีนที่เกิดจากสายโซ่พอลิเปปไทด์หนึ่งสายจะมีลักษณะเป็นก้อน ซึ่งแต่ละส่วนไม่สามารถทำงานและแยกจากกัน โดยคงไว้ซึ่งโครงสร้างเดียวกัน อย่างไรก็ตาม ในบางกรณี ที่มีปริมาณกรดอะมิโนตกค้างสูง (มากกว่า 200) ไม่พบบริเวณกะทัดรัดอิสระเพียงแห่งเดียวของสายพอลิเปปไทด์เดียวในโครงสร้างสามมิติ ชิ้นส่วนเหล่านี้ของสายโพลีเปปไทด์ซึ่งมีสมบัติคล้ายกับโปรตีนทรงกลมอิสระเรียกว่า โมดูล หรือ โดเมน . ตัวอย่างเช่น มีสองโดเมนในดีไฮโดรจีเนส โดเมนหนึ่งจับ NAD + และโดเมนนี้มีโครงสร้างคล้ายกันสำหรับดีไฮโดรจีเนสที่ขึ้นกับ NAD ทั้งหมด และโดเมนอื่นจับซับสเตรตและโครงสร้างที่แตกต่างกันสำหรับดีไฮโดรจีเนสที่ต่างกัน
การสังเคราะห์กรดไขมัน ซึ่งเป็นสายโซ่โพลีเปปไทด์เดี่ยว มี 7 โดเมนเพื่อเร่งปฏิกิริยา 7 ปฏิกิริยา สันนิษฐานว่าครั้งหนึ่งโดเมนสังเคราะห์ถูกรวมเป็นหนึ่งโปรตีนอันเป็นผลมาจากการหลอมรวมของยีน การเชื่อมต่อของโมดูล (โดเมน) เป็นโปรตีนตัวเดียวมีส่วนทำให้เกิดการเกิดขึ้นอย่างรวดเร็วและวิวัฒนาการของโปรตีนที่ใช้งานได้ใหม่
แอคทีฟไซต์ของโปรตีนเป็นที่ยึดเหนี่ยวของโปรตีนกับลิแกนด์ ไซต์ถูกสร้างขึ้นบนพื้นผิวของทรงกลมซึ่งสามารถยึดติดกับโมเลกุลอื่น ๆ ที่เรียกว่า ลิแกนด์ . ศูนย์กลางที่แอคทีฟของโปรตีนเกิดขึ้นจากกลุ่มด้านข้างของกรดอะมิโนที่อยู่ใกล้ระดับโครงสร้างตติยภูมิ ในลำดับเชิงเส้นของสายโซ่เปปไทด์ พวกมันสามารถอยู่ในระยะห่างที่แยกออกจากกันอย่างมีนัยสำคัญ โปรตีนมีความจำเพาะสูงเมื่อทำปฏิกิริยากับแกนด์ ความจำเพาะสูงของการทำงานร่วมกันของโปรตีนกับลิแกนด์นั้นมาจากการเติมเต็มของโครงสร้างของแอคทีฟไซต์ของโปรตีนที่มีโครงสร้างของลิแกนด์ การเติมเต็ม คือความสัมพันธ์เชิงพื้นที่และทางเคมีของโมเลกุลที่มีปฏิสัมพันธ์ ตำแหน่งการจับลิแกนด์ของโปรตีนมักตั้งอยู่ระหว่างโดเมน (ตัวอย่างเช่น ตำแหน่งการจับลิแกนด์ของทริปซินมี 2 โดเมนที่คั่นด้วยร่อง)
การทำงานของโปรตีนขึ้นอยู่กับปฏิสัมพันธ์เฉพาะของพวกมันกับลิแกนด์ โปรตีน 50,000 ตัวที่มีจุดศูนย์กลางเฉพาะที่สามารถจับกับลิแกนด์ที่จำเพาะได้ และเนื่องจากลักษณะโครงสร้างของศูนย์กลางที่ออกฤทธิ์ จึงแสดงหน้าที่โดยธรรมชาติของพวกมัน เห็นได้ชัดว่าโครงสร้างหลักประกอบด้วยข้อมูลเกี่ยวกับหน้าที่ของโปรตีน
โครงสร้างควอเทอร์นารี- นี่คือระดับสูงสุดของการจัดโครงสร้าง ซึ่งเป็นไปไม่ได้สำหรับโปรตีนทั้งหมด โครงสร้างควอเทอร์นารีเป็นที่เข้าใจกันว่าเป็นวิธีการวางสายโพลีเปปไทด์ในอวกาศและการก่อตัวของการก่อรูปโมเลกุลขนาดใหญ่เพียงอันเดียวในด้านโครงสร้างและการทำงาน แต่ละสายโซ่โพลีเปปไทด์เรียกว่า โปรโตเมอร์ หรือ หน่วยย่อย ส่วนใหญ่มักไม่มีกิจกรรมทางชีวภาพ โปรตีนได้รับความสามารถนี้ด้วยวิธีบางอย่างของการเชื่อมโยงเชิงพื้นที่ของโปรโตเมอร์ที่เป็นส่วนประกอบ โมเลกุลที่ได้จะเรียกว่า โอลิโกเมอร์ (มัลติเมอร์) .
โครงสร้างควอเทอร์นารีมีความเสถียรโดยพันธะที่ไม่ใช่โควาเลนต์ที่เกิดขึ้นระหว่างแผ่นสัมผัสของโปรโตเมอร์ซึ่งมีปฏิสัมพันธ์ซึ่งกันและกันตามประเภทของการเติมเต็ม
โปรตีนที่มีโครงสร้างควอเทอร์นารีประกอบด้วยเอ็นไซม์หลายชนิด (แลคเตท ดีไฮโดรจีเนส กลูตาเมตดีไฮโดรจีเนส ฯลฯ) รวมทั้งเฮโมโกลบิน โปรตีนไมโอซินหดตัวของกล้ามเนื้อ โปรตีนบางชนิดมีหน่วยย่อยจำนวนน้อย 2-8 โปรตีนบางชนิดมีหน่วยย่อยเป็นร้อยหรือเป็นพันหน่วย ตัวอย่างเช่น โปรตีนไวรัสโมเสกยาสูบมี 2130 หน่วยย่อย
ตัวอย่างทั่วไปของโปรตีนที่มีโครงสร้างควอเทอร์นารีคือเฮโมโกลบิน โมเลกุลของเฮโมโกลบินประกอบด้วย 4 หน่วยย่อย ได้แก่ สายโซ่โพลีเปปไทด์ซึ่งแต่ละอันเกี่ยวข้องกับ heme ซึ่งสายโซ่โพลีเปปไทด์ 2 สายเรียกว่า -2afla และ -2beta ต่างกันในโครงสร้างหลักและความยาวของสายโซ่โพลีเปปไทด์
พันธะที่สร้างโครงสร้างควอเทอร์นารีมีความแข็งแรงน้อยกว่า ภายใต้อิทธิพลของสารบางชนิด โปรตีนจะถูกแยกออกเป็นหน่วยย่อยที่แยกจากกัน เมื่อเอาสารออก หน่วยย่อยสามารถรวมตัวกันอีกครั้งและฟื้นฟูการทำงานทางชีววิทยาของโปรตีน ดังนั้น เมื่อเติมยูเรียลงในสารละลายเฮโมโกลบิน ยูเรียจะแบ่งออกเป็น 4 หน่วยย่อย เมื่อกำจัดยูเรีย บทบาทโครงสร้างและหน้าที่ของเฮโมโกลบินกลับคืนมา
โครงสร้างโมดูล | ธีม |
หน่วยโมดูลาร์ 1 | 1.1. การจัดระเบียบโครงสร้างของโปรตีน ขั้นตอนของการก่อตัวของรูปแบบดั้งเดิมของโปรตีน 1.2. พื้นฐานของการทำงานของโปรตีน ยาที่เป็นแกนด์ที่มีผลต่อการทำงานของโปรตีน 1.3. การเปลี่ยนสภาพของโปรตีนและความเป็นไปได้ของการฟื้นฟูโดยธรรมชาติ |
หน่วยโมดูลาร์2 | 1.4. คุณสมบัติของโครงสร้างและการทำงานของโปรตีนโอลิโกเมอร์ในตัวอย่างของเฮโมโกลบิน 1.5. รักษารูปแบบดั้งเดิมของโปรตีนในเซลล์ 1.6. โปรตีนหลากหลายชนิด ตระกูลโปรตีนในตัวอย่างของอิมมูโนโกลบูลิน 1.7. คุณสมบัติทางกายภาพและเคมีของโปรตีนและวิธีการแยกออก |
หน่วยโมดูลาร์ 1 การจัดโครงสร้างโปรตีนโมโนเมอร์และพื้นฐานของการทำงาน
วัตถุประสงค์การเรียนรู้ เพื่อให้สามารถ:
1. ใช้ความรู้เกี่ยวกับลักษณะโครงสร้างของโปรตีนและการพึ่งพาการทำงานของโปรตีนในโครงสร้างเพื่อทำความเข้าใจกลไกของการพัฒนาของกรรมพันธุ์และโปรตีนที่ได้มา
2. อธิบายกลไกการรักษาของยาบางชนิดในรูปของลิแกนด์ที่ทำปฏิกิริยากับโปรตีนและเปลี่ยนแปลงกิจกรรมของยา
3. ใช้ความรู้เกี่ยวกับโครงสร้างและความสามารถในการกำหนดโครงสร้างของโปรตีนเพื่อทำความเข้าใจความไม่แน่นอนของโครงสร้างและหน้าที่การใช้งาน และแนวโน้มที่จะเสื่อมสภาพภายใต้สภาวะที่เปลี่ยนแปลง
4. อธิบายการใช้สารทำให้เสียสภาพเป็นวิธีการฆ่าเชื้อวัสดุและอุปกรณ์ทางการแพทย์ รวมทั้งยาฆ่าเชื้อ
รู้:
1. ระดับการจัดโครงสร้างโปรตีน
2. ความสำคัญของโครงสร้างหลักของโปรตีนซึ่งกำหนดความหลากหลายทางโครงสร้างและหน้าที่ของโปรตีน
3. กลไกการก่อตัวของศูนย์แอคทีฟในโปรตีนและปฏิสัมพันธ์เฉพาะกับแกนด์ซึ่งรองรับการทำงานของโปรตีน
4. ตัวอย่างอิทธิพลของลิแกนด์จากภายนอก (ยา สารพิษ สารพิษ) ที่มีต่อโครงสร้างและการทำงานของโปรตีน
5. สาเหตุและผลกระทบของการเสื่อมสภาพของโปรตีน ปัจจัยที่ก่อให้เกิดการเสียสภาพ
6. ตัวอย่างการใช้ปัจจัยที่ทำให้เสียสภาพในยาเป็นน้ำยาฆ่าเชื้อ และวิธีการฆ่าเชื้อเครื่องมือแพทย์
หัวข้อ 1.1. การจัดระเบียบโครงสร้างของโปรตีน ขั้นตอนการสร้างชาวพื้นเมือง
โครงสร้างโปรตีน
โปรตีนเป็นโมเลกุลของพอลิเมอร์ซึ่งมีโมโนเมอร์เพียง 20 α-amino acids ชุดและลำดับการเชื่อมต่อของกรดอะมิโนในโปรตีนถูกกำหนดโดยโครงสร้างของยีนใน DNA ของแต่ละบุคคล โปรตีนแต่ละตัวทำหน้าที่ของตัวเองตามโครงสร้างเฉพาะ ชุดของโปรตีนของสิ่งมีชีวิตที่กำหนดจะกำหนดลักษณะฟีโนไทป์ของมันเช่นเดียวกับการปรากฏตัวของโรคทางพันธุกรรมหรือความโน้มเอียงในการพัฒนาของพวกเขา
1. กรดอะมิโนที่ประกอบเป็นโปรตีน พันธะเปปไทด์โปรตีนเป็นโพลีเมอร์ที่สร้างจากโมโนเมอร์ - กรด α-อะมิโน 20 ตัว ซึ่งมีสูตรทั่วไปคือ
กรดอะมิโนแตกต่างกันในโครงสร้าง ขนาด คุณสมบัติทางเคมีกายภาพของอนุมูลที่ติดอยู่กับอะตอมของ α-คาร์บอน กลุ่มการทำงานของกรดอะมิโนกำหนดคุณสมบัติของคุณสมบัติของกรดα-อะมิโนต่างๆ อนุมูลที่พบในกรด α-amino สามารถแบ่งออกเป็นหลายกลุ่ม:
โพรลีนแตกต่างจากโปรตีนโมโนเมอร์ 19 ชนิดอื่น ๆ ไม่ใช่กรดอะมิโน แต่เป็นกรดอิมิโน เรดิคัลในโพรลีนสัมพันธ์กับทั้งอะตอม α-คาร์บอนและกลุ่มอิมิโน
 กรดอะมิโนมีความสามารถในการละลายในน้ำแตกต่างกันนี่เป็นเพราะความสามารถของอนุมูลอิสระในการทำปฏิกิริยากับน้ำ
กรดอะมิโนมีความสามารถในการละลายในน้ำแตกต่างกันนี่เป็นเพราะความสามารถของอนุมูลอิสระในการทำปฏิกิริยากับน้ำ
ถึง ชอบน้ำรวมถึงอนุมูลที่มีหมู่ฟังก์ชันประจุลบ ประจุบวก และขั้วที่ไม่มีประจุ
ถึง ชอบน้ำรวมถึงอนุมูลที่มีหมู่เมทิล สายโซ่อะลิฟาติกหรือวัฏจักร
2. พันธะเปปไทด์เชื่อมโยงกรดอะมิโนเข้ากับเปปไทด์ในระหว่างการสังเคราะห์เปปไทด์ กลุ่ม α-carboxyl ของกรดอะมิโนหนึ่งตัวจะทำปฏิกิริยากับกลุ่ม α-amino ของกรดอะมิโนอีกตัวหนึ่งเพื่อสร้าง พันธะเปปไทด์:
โปรตีนเป็นโพลีเปปไทด์ กล่าวคือ พอลิเมอร์เชิงเส้นของกรดα-อะมิโนที่เชื่อมต่อด้วยพันธะเปปไทด์ (รูปที่ 1.1.)
 ข้าว. 1.1. คำศัพท์ที่ใช้อธิบายโครงสร้างของเปปไทด์
ข้าว. 1.1. คำศัพท์ที่ใช้อธิบายโครงสร้างของเปปไทด์
โมโนเมอร์ของกรดอะมิโนที่ประกอบเป็นโพลีเปปไทด์เรียกว่า กรดอะมิโนตกค้างห่วงโซ่ของกลุ่มที่ทำซ้ำ - NH-CH-CO- แบบฟอร์ม กระดูกสันหลังเปปไทด์เรซิดิวกรดอะมิโนที่มีหมู่ α-อะมิโนอิสระเรียกว่าปลาย N และตัวที่มีหมู่ α-คาร์บอกซิลอิสระเรียกว่าปลาย C เปปไทด์เขียนและอ่านจากปลาย N ถึงปลาย C
พันธะเปปไทด์ที่เกิดจากกลุ่มอิมิโนของโพรลีนแตกต่างจากพันธะเปปไทด์อื่น: อะตอมไนโตรเจนของกลุ่มเปปไทด์ขาดไฮโดรเจน
แต่มีพันธะกับอนุมูลอิสระ ส่งผลให้ด้านหนึ่งของวัฏจักรรวมอยู่ในกระดูกสันหลังของเปปไทด์:
 เปปไทด์ต่างกันในองค์ประกอบของกรดอะมิโน จำนวนกรดอะมิโนและลำดับของกรดอะมิโน เช่น Ser-Ala-Glu-Gis และ His-Glu-Ala-Ser เป็นเปปไทด์สองชนิดที่แตกต่างกัน
เปปไทด์ต่างกันในองค์ประกอบของกรดอะมิโน จำนวนกรดอะมิโนและลำดับของกรดอะมิโน เช่น Ser-Ala-Glu-Gis และ His-Glu-Ala-Ser เป็นเปปไทด์สองชนิดที่แตกต่างกัน
พันธะเปปไทด์มีความแข็งแรงมาก และจำเป็นต้องมีสภาวะที่รุนแรงสำหรับการไฮโดรไลซิสที่ไม่ใช่เอนไซม์: โปรตีนที่วิเคราะห์จะถูกไฮโดรไลซ์ในกรดไฮโดรคลอริกเข้มข้นที่อุณหภูมิประมาณ 110°C เป็นเวลา 24 ชั่วโมง ในเซลล์ที่มีชีวิต พันธะเปปไทด์สามารถถูกทำลายได้โดย เอนไซม์โปรตีโอไลติก,เรียกว่า โปรตีเอสหรือ เปปไทด์ไฮโดรเลส
3. โครงสร้างหลักของโปรตีนกรดอะมิโนตกค้างในสายโซ่เปปไทด์ของโปรตีนต่างๆ ไม่ได้สลับกันแบบสุ่ม แต่จะจัดเรียงตามลำดับที่แน่นอน ลำดับเชิงเส้นหรือลำดับของเรซิดิวกรดอะมิโนในสายพอลิเปปไทด์เรียกว่า โครงสร้างหลักของโปรตีน
โครงสร้างหลักของโปรตีนแต่ละชนิดถูกเข้ารหัสในโมเลกุลดีเอ็นเอ (ในบริเวณที่เรียกว่ายีน) และนำไปใช้ในระหว่างการถอดรหัส (เขียนข้อมูลใหม่บน mRNA) และการแปล (การสังเคราะห์โครงสร้างหลักของโปรตีน) ดังนั้นโครงสร้างหลักของโปรตีนของแต่ละบุคคลจึงเป็นข้อมูลที่สืบทอดมาจากพ่อแม่สู่ลูกซึ่งกำหนดลักษณะโครงสร้างของโปรตีนของสิ่งมีชีวิตที่กำหนดซึ่งหน้าที่ของโปรตีนที่มีอยู่ขึ้นอยู่กับ (รูปที่ 1.2.)
 ข้าว. 1.2. ความสัมพันธ์ระหว่างจีโนไทป์และโครงสร้างของโปรตีนที่สังเคราะห์ในร่างกายของแต่ละบุคคล
ข้าว. 1.2. ความสัมพันธ์ระหว่างจีโนไทป์และโครงสร้างของโปรตีนที่สังเคราะห์ในร่างกายของแต่ละบุคคล
โปรตีนแต่ละตัวในร่างกายมนุษย์ประมาณ 100,000 ตัวมี มีเอกลักษณ์โครงสร้างหลัก โมเลกุลของโปรตีนชนิดหนึ่ง (เช่น อัลบูมิน) มีการสลับของกรดอะมิโนที่ตกค้างเหมือนกัน ซึ่งแยกอัลบูมินออกจากโปรตีนแต่ละชนิด
ลำดับของเรซิดิวกรดอะมิโนในสายเปปไทด์ถือได้ว่าเป็นรูปแบบของการบันทึกข้อมูล ข้อมูลนี้กำหนดช่องว่างเชิงพื้นที่ของสายเปปไทด์เชิงเส้นให้เป็นโครงสร้างสามมิติที่มีขนาดกะทัดรัดมากขึ้นซึ่งเรียกว่า โครงสร้างกระรอก. กระบวนการของการก่อตัวของโครงสร้างโปรตีนที่ใช้งานได้เรียกว่า พับ.
4. โครงสร้างของโปรตีนการหมุนอิสระในกระดูกสันหลังของเปปไทด์เป็นไปได้ระหว่างอะตอมไนโตรเจนของกลุ่มเปปไทด์กับอะตอม α-คาร์บอนที่อยู่ใกล้เคียง รวมทั้งระหว่างอะตอมของคาร์บอนิลกับคาร์บอนิลกลุ่มคาร์บอนิล เนื่องจากการทำงานร่วมกันของหมู่ฟังก์ชันของกรดอะมิโนตกค้าง โครงสร้างหลักของโปรตีนจึงสามารถได้รับโครงสร้างเชิงพื้นที่ที่ซับซ้อนมากขึ้น ในโปรตีนทรงกลมการพับสองระดับหลักของโครงสร้างของเปปไทด์มีความโดดเด่น: รองและ โครงสร้างระดับอุดมศึกษา
โครงสร้างรองของโปรตีน- นี่คือโครงสร้างเชิงพื้นที่ที่เกิดขึ้นจากการก่อตัวของพันธะไฮโดรเจนระหว่างหมู่ฟังก์ชัน -C=O และ -NH- ของแกนหลักของเปปไทด์ ในกรณีนี้ ห่วงโซ่เปปไทด์สามารถรับโครงสร้างปกติได้สองประเภท: α-helicesและ โครงสร้าง β
ที่ α-helicesพันธะไฮโดรเจนเกิดขึ้นระหว่างอะตอมออกซิเจนของกลุ่มคาร์บอนิลและไฮโดรเจนของอะไมด์ไนโตรเจนของกรดอะมิโนที่ 4 จากนั้น สายข้างของกรดอะมิโนตกค้าง
ตั้งอยู่รอบนอกของเกลียวไม่มีส่วนร่วมในการก่อตัวของโครงสร้างรอง (รูปที่ 1.3.)
อนุมูลขนาดใหญ่หรืออนุมูลที่มีประจุเดียวกันป้องกันการก่อตัวของ α-helix สารตกค้างของโพรลีนซึ่งมีโครงสร้างเป็นวงแหวนจะขัดขวาง α-helix เนื่องจากการขาดไฮโดรเจนที่อะตอมไนโตรเจนในสายโซ่เปปไทด์จึงเป็นไปไม่ได้ที่จะสร้างพันธะไฮโดรเจน พันธะระหว่างไนโตรเจนและอะตอม α-คาร์บอนเป็นส่วนหนึ่งของวัฏจักรโพรลีน ดังนั้นกระดูกสันหลังของเปปไทด์จึงเกิดการโค้งงอในที่นี้
β-โครงสร้างเกิดขึ้นระหว่างบริเวณเส้นตรงของแกนหลักของเปปไทด์ของสายพอลิเปปไทด์หนึ่งสาย ทำให้เกิดโครงสร้างที่พับ สายโพลีเปปไทด์หรือส่วนของพวกมันสามารถก่อรูปได้ ขนานหรือ โครงสร้าง β ที่ต้านขนานกันในกรณีแรก ขั้ว N- และ C ของสายเปปไทด์ที่มีปฏิสัมพันธ์ตรงกัน และในกรณีที่สอง ขั้วทั้งสองมีทิศทางตรงกันข้าม (รูปที่ 1.4)
ข้าว. 1.3. โครงสร้างรองของโปรตีน - α-helix
 ข้าว. 1.4. โครงสร้างจีบแบบขนานและแบบคู่ขนาน
ข้าว. 1.4. โครงสร้างจีบแบบขนานและแบบคู่ขนาน
โครงสร้าง β ถูกระบุด้วยลูกศรกว้าง: A - โครงสร้างต้านขนาน β B - โครงสร้างจีบแบบคู่ขนาน
ในโปรตีนบางชนิด โครงสร้าง β สามารถเกิดขึ้นได้เนื่องจากการก่อตัวของพันธะไฮโดรเจนระหว่างอะตอมของแกนหลักเปปไทด์ของสายโซ่โพลีเปปไทด์ที่ต่างกัน
ยังพบในโปรตีน พื้นที่ที่มีระดับทุติยภูมิผิดปกติโครงสร้างซึ่งรวมถึงส่วนโค้ง ลูป การหมุนของแกนหลักโพลีเปปไทด์ พวกมันมักจะอยู่ในสถานที่ที่ทิศทางของสายโซ่เปปไทด์เปลี่ยนไป ตัวอย่างเช่น ในระหว่างการก่อตัวของโครงสร้างแผ่น β ขนาน
ด้วยการมีอยู่ของ α-helices และ β-โครงสร้าง โปรตีนทรงกลมสามารถแบ่งออกเป็นสี่ประเภท
ข้าว. 1.5. โครงสร้างรองของ myoglobin (A) และ hemoglobin β-chain (B) ซึ่งประกอบด้วย α-helices แปดตัว

 ข้าว. 1.6. โครงสร้างรองของไอโซเมอเรสไตรโอสฟอสเฟตและโดเมนไคเนสไพรูเวต
ข้าว. 1.6. โครงสร้างรองของไอโซเมอเรสไตรโอสฟอสเฟตและโดเมนไคเนสไพรูเวต
 ข้าว. 1.7. โครงสร้างรองของโดเมนคงที่อิมมูโนโกลบูลิน (A) และเอนไซม์ซูเปอร์ออกไซด์ดิสมิวเตส (B)
ข้าว. 1.7. โครงสร้างรองของโดเมนคงที่อิมมูโนโกลบูลิน (A) และเอนไซม์ซูเปอร์ออกไซด์ดิสมิวเตส (B)
ที่ หมวดที่สี่รวมโปรตีนที่มีโครงสร้างรองปกติจำนวนเล็กน้อยในองค์ประกอบ โปรตีนเหล่านี้รวมถึงโปรตีนขนาดเล็กที่อุดมด้วยซิสเทอีนหรือเมทัลโลโปรตีน
โครงสร้างตติยภูมิของโปรตีน- ประเภทของโครงสร้างที่เกิดขึ้นจากการทำงานร่วมกันระหว่างอนุมูลของกรดอะมิโนซึ่งสามารถอยู่ในระยะที่ห่างจากกันมากในสายเปปไทด์ ในกรณีนี้ โปรตีนส่วนใหญ่สร้างโครงสร้างเชิงพื้นที่คล้ายกับทรงกลม (โปรตีนทรงกลม)
เนื่องจากอนุมูลที่ไม่ชอบน้ำของกรดอะมิโนมักจะรวมกันด้วยความช่วยเหลือของสิ่งที่เรียกว่า ปฏิกิริยาที่ไม่ชอบน้ำและแรงระหว่างโมเลกุลของ van der Waals แกนกลางที่ไม่ชอบน้ำหนาแน่นจะก่อตัวขึ้นภายในโปรตีนทรงกลม อนุมูลอิสระที่ชอบน้ำและไม่แตกตัวเป็นไอออนส่วนใหญ่ตั้งอยู่บนผิวของโปรตีนและกำหนดความสามารถในการละลายในน้ำ
 ข้าว. 1.8. ประเภทของพันธะที่เกิดขึ้นระหว่างอนุมูลของกรดอะมิโนระหว่างการก่อตัวของโครงสร้างระดับอุดมศึกษาของโปรตีน
ข้าว. 1.8. ประเภทของพันธะที่เกิดขึ้นระหว่างอนุมูลของกรดอะมิโนระหว่างการก่อตัวของโครงสร้างระดับอุดมศึกษาของโปรตีน
1 - พันธะไอออนิก- เกิดขึ้นระหว่างกลุ่มฟังก์ชันที่มีประจุบวกและลบ
2 - พันธะไฮโดรเจน- เกิดขึ้นระหว่าง hydrophilic uncharged และ hydrophilic group อื่น ๆ
3 - ปฏิกิริยาที่ไม่ชอบน้ำ- เกิดขึ้นระหว่างอนุมูลที่ไม่ชอบน้ำ
4 - พันธะซัลไฟด์- เกิดขึ้นจากการเกิดออกซิเดชันของกลุ่ม SH ของซิสเทอีนตกค้างและปฏิสัมพันธ์ซึ่งกันและกัน
สารตกค้างของกรดอะมิโนที่ชอบน้ำภายในแกนที่ไม่ชอบน้ำสามารถโต้ตอบกันได้โดยใช้ ไอออนิกและ พันธะไฮโดรเจน(รูปที่ 1.8).
พันธะไอออนิกและไฮโดรเจน เช่นเดียวกับปฏิกิริยาที่ไม่ชอบน้ำ เป็นหนึ่งในพันธะที่อ่อนแอ: พลังงานของพวกมันนั้นมากกว่าพลังงานของการเคลื่อนที่ด้วยความร้อนของโมเลกุลที่อุณหภูมิห้องเล็กน้อย โครงสร้างของโปรตีนจะคงอยู่โดยการเกิดพันธะที่อ่อนแอดังกล่าวจำนวนมาก เนื่องจากอะตอมที่ประกอบเป็นโปรตีนมีการเคลื่อนที่อย่างต่อเนื่อง จึงเป็นไปได้ที่จะทำลายพันธะที่อ่อนแอและก่อตัวเป็นพันธะอื่นๆ ซึ่งนำไปสู่การเคลื่อนไหวเล็กๆ ของแต่ละส่วนของสายโซ่โพลีเปปไทด์ คุณสมบัติของโปรตีนนี้ในการเปลี่ยนรูปแบบอันเป็นผลมาจากการทำลายบางส่วนและสร้างพันธะที่อ่อนแออื่น ๆ เรียกว่า lability เชิงโครงสร้าง
ร่างกายมนุษย์มีระบบที่รองรับ สภาวะสมดุล- ความคงตัวของสภาพแวดล้อมภายในภายในขอบเขตที่ยอมรับได้สำหรับสิ่งมีชีวิตที่มีสุขภาพดี ภายใต้สภาวะของสภาวะสมดุล การเปลี่ยนแปลงเล็กน้อยในโครงสร้างจะไม่รบกวนโครงสร้างและหน้าที่โดยรวมของโปรตีน โครงสร้างที่ออกฤทธิ์ตามหน้าที่ของโปรตีนเรียกว่า รูปแบบพื้นเมืองการเปลี่ยนแปลงในสภาพแวดล้อมภายใน (เช่น ความเข้มข้นของกลูโคส ไอออนของ Ca โปรตอน ฯลฯ) นำไปสู่การเปลี่ยนแปลงในรูปแบบและการหยุดชะงักของการทำงานของโปรตีน
โครงสร้างระดับอุดมศึกษาของโปรตีนบางชนิดมีความเสถียร พันธะซัลไฟด์,เกิดขึ้นจากอันตรกิริยาของหมู่ -SH ของสองเรซิดิว
 ข้าว. 1.9. การก่อตัวของพันธะไดซัลไฟด์ในโมเลกุลโปรตีน
ข้าว. 1.9. การก่อตัวของพันธะไดซัลไฟด์ในโมเลกุลโปรตีน
ซิสเทอีน (รูปที่ 1.9) โปรตีนภายในเซลล์ส่วนใหญ่ไม่มีพันธะโควาเลนต์ไดซัลไฟด์ในโครงสร้างระดับอุดมศึกษา การปรากฏตัวของพวกมันเป็นลักษณะของโปรตีนที่หลั่งออกมาจากเซลล์ ซึ่งทำให้มั่นใจในความเสถียรที่มากขึ้นในสภาวะนอกเซลล์ ดังนั้นพันธะซัลไฟด์จึงมีอยู่ในโมเลกุลของอินซูลินและอิมมูโนโกลบูลิน
อินซูลิน- ฮอร์โมนโปรตีนที่สังเคราะห์ขึ้นในเซลล์ β ของตับอ่อนและหลั่งเข้าสู่กระแสเลือดเพื่อตอบสนองต่อการเพิ่มความเข้มข้นของกลูโคสในเลือด ในโครงสร้างของอินซูลิน มีพันธะไดซัลไฟด์สองพันธะที่เชื่อมต่อสายโซ่ A และ B ของโพลีเปปไทด์ และพันธะไดซัลไฟด์หนึ่งพันธะภายในสาย A (รูปที่ 1.10)
 ข้าว. 1.10. พันธะซัลไฟด์ในโครงสร้างของอินซูลิน
ข้าว. 1.10. พันธะซัลไฟด์ในโครงสร้างของอินซูลิน
5. โครงสร้างรองสุดยอดของโปรตีนในโปรตีนที่แตกต่างกันในโครงสร้างและหน้าที่หลัก บางครั้ง การรวมกันและการวางซ้อนของโครงสร้างทุติยภูมิที่คล้ายคลึงกันซึ่งเรียกว่าโครงสร้าง supersecondary มันครองตำแหน่งกลางระหว่างโครงสร้างทุติยภูมิและตติยภูมิเนื่องจากเป็นส่วนผสมเฉพาะขององค์ประกอบโครงสร้างทุติยภูมิในระหว่างการก่อตัวของโครงสร้างระดับอุดมศึกษาของโปรตีน โครงสร้าง supersecondary มีชื่อเฉพาะเช่น "α-helix-turn-a-helix", "leucine zipper", "zinc finger" เป็นต้น โครงสร้าง supersecondary ดังกล่าวเป็นลักษณะของโปรตีนที่จับกับดีเอ็นเอ
"ลิวซีนซิป".โครงสร้างซูเปอร์ทุติยภูมิประเภทนี้ใช้เชื่อมต่อโปรตีนสองชนิด บนพื้นผิวของโปรตีนที่มีปฏิสัมพันธ์กันมีบริเวณลาน α-helical ที่มีลิวซีนตกค้างอย่างน้อยสี่ตัว ลิวซีนตกค้างใน α-helix มีกรดอะมิโน 6 ตัวแยกจากกัน เนื่องจากแต่ละรอบของ α-helix มีเรซิดิวกรดอะมิโน 3.6 ตัว จึงพบอนุมูลลิวซีนบนผิวของเทิร์นอื่นๆ ลิวซีนตกค้างของ α-helix ของโปรตีนหนึ่งสามารถโต้ตอบกับลิวซีนตกค้างของโปรตีนอื่น (ปฏิกิริยาไม่ชอบน้ำ) ซึ่งเชื่อมต่อเข้าด้วยกัน (รูปที่ 1.11) โปรตีนที่จับกับดีเอ็นเอจำนวนมากทำงานเป็นส่วนหนึ่งของสารเชิงซ้อนโอลิโกเมอริก ซึ่งแต่ละหน่วยย่อยจะเชื่อมโยงถึงกันโดย "ลิวซีนซิป"
 ข้าว. 1.11. "ลิวซีนซิป" ระหว่างบริเวณ α-helical ของโปรตีนสองตัว
ข้าว. 1.11. "ลิวซีนซิป" ระหว่างบริเวณ α-helical ของโปรตีนสองตัว
ฮิสโตนเป็นตัวอย่างของโปรตีนดังกล่าว ฮิสโตนส์- โปรตีนนิวเคลียร์ซึ่งรวมถึงกรดอะมิโนที่มีประจุบวกจำนวนมาก - อาร์จินีนและไลซีน (มากถึง 80%) โมเลกุลของฮิสโตนถูกรวมเข้ากับสารเชิงซ้อนโอลิโกเมอร์ที่มีโมโนเมอร์แปดตัวโดยใช้ "สารยึดลิวซีน" แม้ว่าจะมีประจุที่มีนัยสำคัญของโมเลกุลเหล่านี้ก็ตาม
"นิ้วสังกะสี"- ความแตกต่างของโครงสร้าง supersecondary ซึ่งเป็นลักษณะของโปรตีนที่จับกับ DNA มีรูปแบบของชิ้นส่วนที่ยืดออกบนพื้นผิวของโปรตีนและมีกรดอะมิโนตกค้างประมาณ 20 ชนิด (รูปที่ 1.12) รูปร่างของ "นิ้วที่เหยียด" ได้รับการสนับสนุนโดยอะตอมของสังกะสีที่เกี่ยวข้องกับอนุมูลของกรดอะมิโนสี่ตัว - ซิสเทอีนตกค้างสองตัวและฮิสทิดีนสองตัว ในบางกรณี แทนที่จะเป็นสารตกค้างของฮิสทิดีน มีซิสเทอีนตกค้างอยู่ ซิสเทอีนตกค้าง 2 ตัวที่เว้นระยะห่างอย่างใกล้ชิดนั้นแยกออกจากส่วนที่เหลือของ Gisili อีก 2 ตัวโดยลำดับ Cys ของกรดอะมิโนประมาณ 12 ตัว บริเวณนี้ของโปรตีนก่อตัวเป็นเกลียว α ซึ่งอนุมูลซึ่งสามารถจับกับบริเวณควบคุมของร่องหลักของ DNA ได้โดยเฉพาะ ความจำเพาะของการผูกมัดของแต่ละบุคคล
 ข้าว. 1.12. โครงสร้างหลักของส่วนของโปรตีนที่จับกับดีเอ็นเอที่สร้างโครงสร้าง "นิ้วสังกะสี" (ตัวอักษรระบุถึงกรดอะมิโนที่ประกอบเป็นโครงสร้างนี้)
ข้าว. 1.12. โครงสร้างหลักของส่วนของโปรตีนที่จับกับดีเอ็นเอที่สร้างโครงสร้าง "นิ้วสังกะสี" (ตัวอักษรระบุถึงกรดอะมิโนที่ประกอบเป็นโครงสร้างนี้)
โปรตีนที่จับกับดีเอ็นเอควบคุมจะขึ้นอยู่กับลำดับของกรดอะมิโนตกค้างที่อยู่ใน "นิ้วสังกะสี" โดยเฉพาะอย่างยิ่ง โครงสร้างดังกล่าวประกอบด้วยตัวรับฮอร์โมนสเตียรอยด์ที่เกี่ยวข้องกับการควบคุมการถอดรหัส (การอ่านข้อมูลจาก DNA ไปยัง RNA)
หัวข้อ 1.2. พื้นฐานของการทำงานของโปรตีน ยาที่เป็นแกนด์ที่มีผลต่อการทำงานของโปรตีน
1. ศูนย์กลางของโปรตีนและการมีปฏิสัมพันธ์กับแกนด์ในระหว่างการก่อตัวของโครงสร้างระดับอุดมศึกษา บนพื้นผิวของโปรตีนที่ใช้งานได้ซึ่งมักจะอยู่ในช่อง ไซต์จะถูกสร้างขึ้นโดยอนุมูลของกรดอะมิโนที่อยู่ห่างไกลกันในโครงสร้างหลัก ไซต์นี้ซึ่งมีโครงสร้างเฉพาะสำหรับโปรตีนที่กำหนดและสามารถโต้ตอบอย่างจำเพาะกับโมเลกุลบางตัวหรือกลุ่มของโมเลกุลที่คล้ายคลึงกัน เรียกว่าไซต์การจับโปรตีนที่มีลิแกนด์หรือไซต์ที่ทำงานอยู่ แกนด์เป็นโมเลกุลที่ทำปฏิกิริยากับโปรตีน
ความจำเพาะสูงปฏิสัมพันธ์ของโปรตีนกับลิแกนด์ได้รับการประกันโดยความสมบูรณ์ของโครงสร้างของศูนย์แอคทีฟที่มีโครงสร้างของลิแกนด์
การเติมเต็มคือความสัมพันธ์เชิงพื้นที่และทางเคมีของพื้นผิวที่มีปฏิสัมพันธ์กัน ศูนย์แอคทีฟจะต้องไม่สัมพันธ์เชิงพื้นที่กับลิแกนด์ที่รวมอยู่ในนั้นเท่านั้น แต่พันธะ (อันตรกิริยาของไอออนิก ไฮโดรเจน และไม่ชอบน้ำ) จะต้องก่อตัวขึ้นระหว่างกลุ่มการทำงานของอนุมูลที่รวมอยู่ในศูนย์กลางแอคทีฟและลิแกนด์ ซึ่งทำให้ลิแกนด์อยู่ใน ศูนย์แอคทีฟ (รูปที่ 1.13 )
 ข้าว. 1.13. ปฏิกิริยาเสริมของโปรตีนกับลิแกนด์
ข้าว. 1.13. ปฏิกิริยาเสริมของโปรตีนกับลิแกนด์
ลิแกนด์บางตัวเมื่อยึดติดกับศูนย์กลางของโปรตีน จะมีบทบาทสำคัญในการทำงานของโปรตีน ลิแกนด์ดังกล่าวเรียกว่าโคแฟคเตอร์และโปรตีนที่มีองค์ประกอบที่ไม่ใช่โปรตีนเรียกว่า โปรตีนที่ซับซ้อน(ตรงกันข้ามกับโปรตีนอย่างง่าย ประกอบด้วยส่วนโปรตีนเท่านั้น) ส่วนที่ไม่ใช่โปรตีนที่ยึดติดกับโปรตีนอย่างแน่นหนาเรียกว่า กลุ่มเทียมตัวอย่างเช่น องค์ประกอบของ myoglobin, hemoglobin และ cytochromes มีกลุ่มเทียมที่ติดอยู่กับศูนย์ที่ใช้งานอยู่อย่างแน่นหนา - heme ที่มีไอออนเหล็ก โปรตีนเชิงซ้อนที่มี heme เรียกว่า hemoproteins
เมื่อลิแกนด์จำเพาะต่อโปรตีน หน้าที่ของโปรตีนเหล่านี้จะแสดงออกมา ดังนั้นอัลบูมินซึ่งเป็นโปรตีนที่สำคัญที่สุดในพลาสมาจึงแสดงหน้าที่การลำเลียงโดยยึดแกนด์ที่ไม่ชอบน้ำเข้ากับศูนย์กลางที่ทำงานอยู่ เช่น กรดไขมัน บิลิรูบิน ยาบางชนิด เป็นต้น (รูปที่ 1.14)
ลิแกนด์ที่ทำปฏิกิริยากับโครงสร้างสามมิติของสายเปปไทด์ไม่เพียงแต่เป็นโมเลกุลอินทรีย์และอนินทรีย์ที่มีน้ำหนักโมเลกุลต่ำเท่านั้น แต่ยังรวมถึงโมเลกุลขนาดใหญ่ด้วย:
ดีเอ็นเอ (ตัวอย่างที่อภิปรายข้างต้นด้วยโปรตีนที่จับกับดีเอ็นเอ);
โพลีแซ็กคาไรด์;
 ข้าว. 1.14. ความสัมพันธ์ระหว่างจีโนไทป์และฟีโนไทป์
ข้าว. 1.14. ความสัมพันธ์ระหว่างจีโนไทป์และฟีโนไทป์
โครงสร้างหลักอันเป็นเอกลักษณ์ของโปรตีนของมนุษย์ ซึ่งเข้ารหัสไว้ในโมเลกุลดีเอ็นเอ ถูกทำให้เป็นจริงในเซลล์ในรูปแบบของโครงสร้างที่มีลักษณะเฉพาะ โครงสร้างแอคทีฟไซต์ และหน้าที่ของโปรตีน
ในกรณีเหล่านี้ โปรตีนรับรู้บริเวณจำเพาะของลิแกนด์ที่สมส่วนกับตำแหน่งการจับ ดังนั้นบนพื้นผิวของเซลล์ตับจึงมีโปรตีนตัวรับสำหรับฮอร์โมนอินซูลินซึ่งมีโครงสร้างโปรตีนด้วย ปฏิสัมพันธ์ของอินซูลินกับตัวรับทำให้เกิดการเปลี่ยนแปลงในรูปแบบและการกระตุ้นระบบสัญญาณซึ่งนำไปสู่การสะสมของสารอาหารในเซลล์ตับหลังรับประทานอาหาร
ดังนั้น, การทำงานของโปรตีนขึ้นอยู่กับปฏิสัมพันธ์เฉพาะของศูนย์กลางการทำงานของโปรตีนกับแกนด์
2. โครงสร้างโดเมนและบทบาทในการทำงานของโปรตีนสายโพลีเปปไทด์แบบยาวของโปรตีนทรงกลมมักจะพับเป็นบริเวณที่มีขนาดกะทัดรัดและค่อนข้างอิสระหลายแห่ง พวกมันมีโครงสร้างตติยภูมิอิสระ คล้ายกับโปรตีนทรงกลม และเรียกว่า โดเมนเนื่องจากโครงสร้างโดเมนของโปรตีน โครงสร้างระดับตติยภูมิจึงง่ายต่อการสร้าง
ในโปรตีนโดเมน ตำแหน่งการจับลิแกนด์มักตั้งอยู่ระหว่างโดเมน ดังนั้นทริปซินจึงเป็นเอนไซม์โปรตีโอไลติกที่ผลิตโดยตับอ่อนส่วนนอกและจำเป็นสำหรับการย่อยโปรตีนในอาหาร มีโครงสร้างแบบสองโดเมน และตำแหน่งจับของทริปซินกับลิแกนด์ - โปรตีนจากอาหาร - ตั้งอยู่ในร่องระหว่างสองโดเมน ในศูนย์ปฏิบัติการ สภาวะที่จำเป็นสำหรับการจับอย่างมีประสิทธิภาพของตำแหน่งเฉพาะของโปรตีนในอาหารและการไฮโดรไลซิสของพันธะเปปไทด์จะถูกสร้างขึ้น
โดเมนที่แตกต่างกันในโปรตีนสามารถเคลื่อนที่สัมพันธ์กันเมื่อศูนย์กลางที่ทำงานโต้ตอบกับลิแกนด์ (รูปที่ 1.15)
เฮกโซคินาเสะ- เอ็นไซม์ที่เร่งปฏิกิริยาฟอสโฟรีเลชั่นของกลูโคสด้วยความช่วยเหลือของเอทีพี แอคทีฟไซต์ของเอ็นไซม์อยู่ในรอยแยกระหว่างสองโดเมน เมื่อเฮกโซไคเนสจับกับกลูโคส โดเมนโดยรอบจะปิดและซับสเตรตจะถูกดักจับ ซึ่งเกิดฟอสโฟรีเลชัน (ดูรูปที่ 1.15)
 ข้าว. 1.15. การจับโดเมนเฮกโซไคเนสกับกลูโคส
ข้าว. 1.15. การจับโดเมนเฮกโซไคเนสกับกลูโคส
ในโปรตีนบางชนิด โดเมนทำหน้าที่อิสระโดยจับกับลิแกนด์ต่างๆ โปรตีนดังกล่าวเรียกว่ามัลติฟังก์ชั่น
3. ยา-ลิแกนด์ที่ส่งผลต่อการทำงานของโปรตีนปฏิสัมพันธ์ของโปรตีนกับลิแกนด์มีความเฉพาะเจาะจง อย่างไรก็ตาม เนื่องจากความสามารถในการกำหนดโครงสร้างของโปรตีนและตำแหน่งแอคทีฟของมัน จึงเป็นไปได้ที่จะเลือกสารอื่นที่สามารถโต้ตอบกับโปรตีนในบริเวณที่ทำงานหรือส่วนอื่นของโมเลกุลได้
สารที่มีโครงสร้างคล้ายกับลิแกนด์ตามธรรมชาติเรียกว่า อะนาล็อกโครงสร้างของลิแกนด์หรือแกนด์ที่ผิดธรรมชาติ นอกจากนี้ยังทำปฏิกิริยากับโปรตีนในบริเวณที่ทำงานอยู่ โครงสร้างแอนะล็อกของลิแกนด์สามารถเพิ่มการทำงานของโปรตีนได้ (ตัวเอก)แล้วก็ลด (ศัตรู).ลิแกนด์และโครงสร้างที่คล้ายคลึงกันจะแข่งขันกันเพื่อจับโปรตีนที่ตำแหน่งเดียวกัน สารดังกล่าวเรียกว่า โมดูเลเตอร์การแข่งขัน(ตัวควบคุม) ของการทำงานของโปรตีน ยาหลายชนิดทำหน้าที่เป็นตัวยับยั้งโปรตีน บางส่วนได้มาจากการดัดแปลงทางเคมีของแกนด์ตามธรรมชาติ สารยับยั้งการทำงานของโปรตีนอาจเป็นยาและสารพิษ
Atropine เป็นตัวยับยั้งการแข่งขันของตัวรับ M-cholinergicอะเซทิลโคลีนเป็นสารสื่อประสาทสำหรับส่งผ่านแรงกระตุ้นของเส้นประสาทผ่านไซแนปส์ cholinergic ในการกระตุ้นให้เกิดการกระตุ้น acetylcholine ที่ปล่อยออกมาในช่อง synaptic จะต้องโต้ตอบกับโปรตีน - ตัวรับของเยื่อหุ้มเซลล์ postsynaptic พบสองประเภท ตัวรับ cholinergic:
ตัวรับ Mนอกจาก acetylcholine แล้วยังเลือกโต้ตอบกับ muscarine (fly agaric toxin) M - ตัวรับ cholinergic มีอยู่ในกล้ามเนื้อเรียบและเมื่อมีปฏิสัมพันธ์กับ acetylcholine จะทำให้เกิดการหดตัว
ตัวรับ Hผูกติดกับนิโคตินโดยเฉพาะ ตัวรับ N-cholinergic พบได้ในประสาทของกล้ามเนื้อโครงร่างลาย
สารยับยั้งจำเพาะ ตัวรับ M-cholinergicคืออะโทรพีน พบในพืชจำพวกเบลลาดอนน่าและเฮนเบน
 Atropine มีกลุ่มการทำงานและการจัดเรียงเชิงพื้นที่คล้ายกับ acetylcholine ในโครงสร้าง ดังนั้นจึงเป็นของตัวยับยั้งการแข่งขันของตัวรับ M-cholinergic เนื่องจากการจับกันของ acetylcholine กับตัวรับ M-cholinergic ทำให้เกิดการหดตัวของกล้ามเนื้อเรียบ atropine จึงถูกใช้เป็นยาที่ช่วยบรรเทาอาการกระตุก (ต้านอาการกระสับกระส่าย).ดังนั้นจึงเป็นที่ทราบกันดีว่าการใช้ atropine เพื่อผ่อนคลายกล้ามเนื้อตาเมื่อดูอวัยวะและบรรเทาอาการกระตุกในอาการจุกเสียดในทางเดินอาหาร ตัวรับ M-cholinergic ยังมีอยู่ที่ส่วนกลาง ระบบประสาท(CNS) ดังนั้นการให้อะโทรปีนในปริมาณมากจึงทำให้เกิด ปฏิกิริยาที่ไม่ต้องการจากด้านข้างของระบบประสาทส่วนกลาง: ความปั่นป่วนของมอเตอร์และจิตใจ, ภาพหลอน, อาการชัก
Atropine มีกลุ่มการทำงานและการจัดเรียงเชิงพื้นที่คล้ายกับ acetylcholine ในโครงสร้าง ดังนั้นจึงเป็นของตัวยับยั้งการแข่งขันของตัวรับ M-cholinergic เนื่องจากการจับกันของ acetylcholine กับตัวรับ M-cholinergic ทำให้เกิดการหดตัวของกล้ามเนื้อเรียบ atropine จึงถูกใช้เป็นยาที่ช่วยบรรเทาอาการกระตุก (ต้านอาการกระสับกระส่าย).ดังนั้นจึงเป็นที่ทราบกันดีว่าการใช้ atropine เพื่อผ่อนคลายกล้ามเนื้อตาเมื่อดูอวัยวะและบรรเทาอาการกระตุกในอาการจุกเสียดในทางเดินอาหาร ตัวรับ M-cholinergic ยังมีอยู่ที่ส่วนกลาง ระบบประสาท(CNS) ดังนั้นการให้อะโทรปีนในปริมาณมากจึงทำให้เกิด ปฏิกิริยาที่ไม่ต้องการจากด้านข้างของระบบประสาทส่วนกลาง: ความปั่นป่วนของมอเตอร์และจิตใจ, ภาพหลอน, อาการชัก
Ditilin เป็นตัวเร่งการแข่งขันของตัวรับ H-cholinergic ที่ยับยั้งการทำงานของประสาทและกล้ามเนื้อ synapses
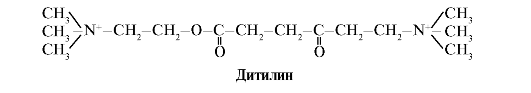 ประสาทและกล้ามเนื้อของกล้ามเนื้อโครงร่างประกอบด้วยตัวรับ H-cholinergic การมีปฏิสัมพันธ์กับ acetylcholine ทำให้กล้ามเนื้อหดตัว ในการผ่าตัดบางอย่างเช่นเดียวกับในการศึกษาส่องกล้อง ใช้ยาที่ทำให้กล้ามเนื้อโครงร่างผ่อนคลาย (ยาคลายกล้ามเนื้อ).ซึ่งรวมถึงไดไทลินซึ่งเป็นอะซิติลโคลีนที่มีโครงสร้างคล้ายคลึงกัน มันยึดติดกับตัวรับ H-cholinergic แต่ต่างจาก acetylcholine เอนไซม์ acetylcholinesterase ถูกทำลายอย่างช้าๆ อันเป็นผลมาจากการเปิดช่องไอออนเป็นเวลานานและการสลับขั้วของเมมเบรนอย่างต่อเนื่อง การนำกระแสประสาทจะหยุดชะงักและเกิดการคลายตัวของกล้ามเนื้อ ในขั้นต้นพบคุณสมบัติเหล่านี้ในพิษของ curare ดังนั้นจึงเรียกว่ายาดังกล่าว คูราริฟอร์ม
ประสาทและกล้ามเนื้อของกล้ามเนื้อโครงร่างประกอบด้วยตัวรับ H-cholinergic การมีปฏิสัมพันธ์กับ acetylcholine ทำให้กล้ามเนื้อหดตัว ในการผ่าตัดบางอย่างเช่นเดียวกับในการศึกษาส่องกล้อง ใช้ยาที่ทำให้กล้ามเนื้อโครงร่างผ่อนคลาย (ยาคลายกล้ามเนื้อ).ซึ่งรวมถึงไดไทลินซึ่งเป็นอะซิติลโคลีนที่มีโครงสร้างคล้ายคลึงกัน มันยึดติดกับตัวรับ H-cholinergic แต่ต่างจาก acetylcholine เอนไซม์ acetylcholinesterase ถูกทำลายอย่างช้าๆ อันเป็นผลมาจากการเปิดช่องไอออนเป็นเวลานานและการสลับขั้วของเมมเบรนอย่างต่อเนื่อง การนำกระแสประสาทจะหยุดชะงักและเกิดการคลายตัวของกล้ามเนื้อ ในขั้นต้นพบคุณสมบัติเหล่านี้ในพิษของ curare ดังนั้นจึงเรียกว่ายาดังกล่าว คูราริฟอร์ม
หัวข้อ 1.3. การสลายตัวของโปรตีนและความเป็นไปได้ของการฟื้นฟูที่เกิดขึ้นเอง
1. เนื่องจากรูปแบบดั้งเดิมของโปรตีนยังคงอยู่เนื่องจากการโต้ตอบที่อ่อนแอ การเปลี่ยนแปลงองค์ประกอบและคุณสมบัติของสภาพแวดล้อมรอบ ๆ โปรตีน ผลกระทบ สารเคมีและปัจจัยทางกายภาพทำให้เกิดการเปลี่ยนแปลงในโครงสร้างของพวกเขา (คุณสมบัติของโครงสร้าง lability) การแตกของพันธะจำนวนมากนำไปสู่การทำลายโครงสร้างดั้งเดิมและการเสื่อมสภาพของโปรตีน
การเสื่อมสภาพของโปรตีน- นี่คือการทำลายโครงสร้างดั้งเดิมของพวกมันภายใต้การกระทำของสารทำให้เสียสภาพซึ่งเกิดจากการแตกของพันธะที่อ่อนแอซึ่งทำให้โครงสร้างเชิงพื้นที่ของโปรตีนเสถียร การเปลี่ยนสภาพจะมาพร้อมกับการทำลายโครงสร้างสามมิติที่เป็นเอกลักษณ์และศูนย์กลางการทำงานของโปรตีนและการสูญเสียกิจกรรมทางชีวภาพ (รูปที่ 1.16)
โมเลกุลที่แปลงสภาพทั้งหมดของโปรตีนหนึ่งตัวจะได้รับโครงสร้างแบบสุ่มที่แตกต่างจากโมเลกุลอื่นของโปรตีนเดียวกัน อนุมูลของกรดอะมิโนที่ก่อตัวเป็นศูนย์แอคทีฟกลับกลายเป็นว่าห่างไกลจากกันในเชิงพื้นที่ กล่าวคือ บริเวณจับเฉพาะของโปรตีนที่มีแกนด์ถูกทำลาย ในระหว่างการแปลงสภาพ โครงสร้างหลักของโปรตีนยังคงไม่เปลี่ยนแปลง
การใช้สารทำให้เสียสภาพในการวิจัยทางชีววิทยาและการแพทย์ในการศึกษาทางชีวเคมี ก่อนการหาสารประกอบที่มีน้ำหนักโมเลกุลต่ำในวัสดุชีวภาพ โปรตีนมักจะถูกกำจัดออกจากสารละลายก่อน ด้วยเหตุนี้จึงมักใช้กรดไตรคลอโรอะซิติก (TCA) หลังจากเติม TCA ลงในสารละลาย โปรตีนที่แปลงสภาพจะตกตะกอนและกรองออกได้ง่าย (ตารางที่ 1.1)
ในทางการแพทย์ สารทำให้เสียสภาพมักใช้ในการฆ่าเชื้อเครื่องมือทางการแพทย์และวัสดุในหม้อนึ่งความดัน (สารทำให้เสียสภาพ - อุณหภูมิสูง) และใช้เป็นน้ำยาฆ่าเชื้อ (แอลกอฮอล์ ฟีนอล คลอรามีน) เพื่อรักษาพื้นผิวที่ปนเปื้อนที่มีจุลินทรีย์ที่ทำให้เกิดโรค
2. การสร้างโปรตีนขึ้นใหม่โดยธรรมชาติ- การพิสูจน์การกำหนดโครงสร้างหลัก โครงสร้าง และหน้าที่ของโปรตีน โปรตีนแต่ละชนิดเป็นผลิตภัณฑ์ของยีนหนึ่งตัวที่มีลำดับกรดอะมิโนเหมือนกันและได้รับโครงสร้างที่เหมือนกันในเซลล์ ข้อสรุปพื้นฐานที่ว่าโครงสร้างหลักของโปรตีนมีข้อมูลเกี่ยวกับโครงสร้างและหน้าที่ของมันอยู่แล้วบนพื้นฐานของความสามารถของโปรตีนบางชนิด (โดยเฉพาะ ribonuclease และ myoglobin) ในการฟื้นสภาพที่เกิดขึ้นเองตามธรรมชาติ - การฟื้นฟูโครงสร้างดั้งเดิมของพวกมันหลังจากการเสียสภาพ
การก่อตัวของโครงสร้างเชิงพื้นที่ของโปรตีนนั้นดำเนินการโดยวิธีการประกอบตัวเอง - กระบวนการที่เกิดขึ้นเองซึ่งโซ่โพลีเปปไทด์ซึ่งมีโครงสร้างหลักที่เป็นเอกลักษณ์ มีแนวโน้มที่จะนำโครงสร้างที่มีพลังงานอิสระต่ำที่สุดในสารละลายมาใช้ ความสามารถในการสร้างโปรตีนใหม่ที่คงโครงสร้างหลักของพวกมันไว้หลังจากการเสียสภาพได้อธิบายไว้ในการทดลองกับเอนไซม์ไรโบนิวคลีเอส
ไรโบนิวคลีเอสเป็นเอนไซม์ที่ทำลายพันธะระหว่างนิวคลีโอไทด์แต่ละตัวในโมเลกุลอาร์เอ็นเอ โปรตีนทรงกลมนี้มีสายโพลีเปปไทด์หนึ่งสาย โครงสร้างระดับอุดมศึกษาซึ่งมีความเสถียรด้วยพันธะซัลไฟด์ที่อ่อนแอจำนวนมากและพันธะสี่พันธะ
การบำบัดไรโบนิวคลีเอสด้วยยูเรียซึ่งทำลายพันธะไฮโดรเจนในโมเลกุล และตัวรีดิวซ์ซึ่งทำลายพันธะไดซัลไฟด์ นำไปสู่การเสื่อมสภาพของเอนไซม์และการสูญเสียกิจกรรม
การกำจัดสารเปลี่ยนสภาพโดยการฟอกไตจะนำไปสู่การฟื้นฟูโครงสร้างและหน้าที่ของโปรตีน กล่าวคือ เพื่อฟื้นคืนชีพ (รูปที่ 1.17).
 ข้าว. 1.17. การเปลี่ยนสภาพและการสร้างใหม่ของไรโบนิวคลีเอส
ข้าว. 1.17. การเปลี่ยนสภาพและการสร้างใหม่ของไรโบนิวคลีเอส
เอ - โครงสร้างดั้งเดิมของไรโบนิวคลีเอสในโครงสร้างตติยภูมิซึ่งมีพันธะซัลไฟด์สี่พันธะ B - โมเลกุลไรโบนิวคลีเอสที่ทำให้เสียสภาพ
B - โมเลกุลไรโบนิวคลีเอสรีเนทีฟพร้อมโครงสร้างและหน้าที่ฟื้นฟู
1. กรอกตาราง 1.2
ตารางที่ 1.2. การจำแนกกรดอะมิโนตามขั้วของอนุมูล
2. เขียนสูตรของเตตราเปปไทด์:
Asp - Pro - เฟิน - Liz
ก) แยกกลุ่มที่เกิดซ้ำในเปปไทด์ที่ก่อรูปกระดูกสันหลังของเปปไทด์และกลุ่มตัวแปรที่แสดงโดยอนุมูลของกรดอะมิโน
b) กำหนด N- และ C-termi;
c) ขีดเส้นใต้พันธะเปปไทด์;
d) เขียนเปปไทด์อื่นที่ประกอบด้วยกรดอะมิโนเดียวกัน
จ) นับจำนวน ตัวเลือก tetrapeptide ที่มีองค์ประกอบของกรดอะมิโนเหมือนกัน
3. อธิบายบทบาทของโครงสร้างหลักของโปรตีนโดยใช้ตัวอย่างการวิเคราะห์เปรียบเทียบฮอร์โมนเปปไทด์ที่มีโครงสร้างคล้ายคลึงกันและวิวัฒนาการอย่างใกล้ชิดสองชนิดของ neurohypophysis ของสัตว์เลี้ยงลูกด้วยนม - ออกซีโตซินและวาโซเพรสซิน (ตารางที่ 1.3)
ตาราง 1.3. โครงสร้างและหน้าที่ของออกซิโทซินและวาโซเพรสซิน
 สำหรับสิ่งนี้:
สำหรับสิ่งนี้:
ก) เปรียบเทียบองค์ประกอบและลำดับกรดอะมิโนของเปปไทด์สองตัว
b) ค้นหาความคล้ายคลึงกันของโครงสร้างหลักของเปปไทด์ทั้งสองและความคล้ายคลึงกันของการกระทำทางชีวภาพของพวกมัน
c) ค้นหาความแตกต่างในโครงสร้างของเปปไทด์ทั้งสองและความแตกต่างในหน้าที่ของพวกมัน
d) ทำการสรุปเกี่ยวกับอิทธิพลของโครงสร้างหลักของเปปไทด์ที่มีต่อหน้าที่ของเปปไทด์
4. อธิบายขั้นตอนหลักในการสร้างโครงสร้างของโปรตีนทรงกลม (โครงสร้างทุติยภูมิ, โครงสร้างตติยภูมิ, แนวคิดของโครงสร้าง supersecondary) ระบุประเภทของพันธะที่เกี่ยวข้องกับการก่อตัวของโครงสร้างโปรตีน อนุมูลของกรดอะมิโนชนิดใดสามารถมีส่วนร่วมในการก่อตัวของปฏิกิริยาที่ไม่ชอบน้ำ พันธะไอออนิก ไฮโดรเจน
ยกตัวอย่าง.
5. กำหนดแนวคิดของ "ความสามารถในการกำหนดรูปแบบโปรตีน" ระบุสาเหตุของการมีอยู่และความสำคัญของโปรตีน
6. อธิบายความหมายของวลีต่อไปนี้: "การทำงานของโปรตีนตามปฏิสัมพันธ์เฉพาะของพวกมันกับลิแกนด์" โดยใช้คำศัพท์และอธิบายความหมายของพวกมัน: โครงสร้างของโปรตีน, แอคทีฟไซต์, ลิแกนด์, คอมพลีเมนต์, ฟังก์ชันโปรตีน
7. ใช้ตัวอย่างใดตัวอย่างหนึ่งเพื่ออธิบายว่าโดเมนคืออะไรและมีบทบาทอย่างไรต่อการทำงานของโปรตีน
งานเพื่อการควบคุมตนเอง
1. ตั้งค่าการแข่งขัน
กลุ่มหน้าที่ในอนุมูลของกรดอะมิโน:
A. กลุ่มคาร์บอกซิล B. กลุ่มไฮดรอกซิล C กลุ่ม Guanidine D. กลุ่มธีออล E. กลุ่มอะมิโน
2. เลือกคำตอบที่ถูกต้อง.
กรดอะมิโนที่มีขั้วไม่มีประจุคือ:
A. Tsis B. Asn
บี กลู จี สาม
3. เลือกคำตอบที่ถูกต้อง.
อนุมูลของกรดอะมิโน:
ก. จัดให้มีความจำเพาะของโครงสร้างหลัก ข. มีส่วนร่วมในการก่อตัวของโครงสร้างตติยภูมิ
B. ตั้งอยู่บนพื้นผิวของโปรตีน พวกมันส่งผลต่อความสามารถในการละลายของโปรตีน D. สร้างศูนย์กลางที่แอคทีฟ
D. มีส่วนร่วมในการก่อตัวของพันธะเปปไทด์
4. เลือกคำตอบที่ถูกต้อง.
ปฏิกิริยาที่ชอบน้ำสามารถเกิดขึ้นได้ระหว่างอนุมูลของกรดอะมิโน:
A. Tre Lay B. Pro สาม
B. Met Ile G. Tir Ala D. Val Fen
5. เลือกคำตอบที่ถูกต้อง.
พันธะไอออนิกสามารถเกิดขึ้นได้ระหว่างอนุมูลของกรดอะมิโน:
A. Gln Asp B. เมษายน Liz
B. Liz Glu G. Geese Asp D. Asn Apr
6. เลือกคำตอบที่ถูกต้อง.
พันธะไฮโดรเจนสามารถเกิดขึ้นได้ระหว่างอนุมูลของกรดอะมิโน:
A. Ser Gln B. Cis Tre
B. Asp Liz G. Glu Asp D. Asn Tre
7. ตั้งค่าการแข่งขัน
ประเภทของพันธะที่เกี่ยวข้องกับการก่อตัวของโครงสร้างโปรตีน:
ก. โครงสร้างปฐมภูมิ ข. โครงสร้างรอง
ข. โครงสร้างระดับตติยภูมิ
ง. โครงสร้าง supersecondary E. โครงสร้าง.
1. พันธะไฮโดรเจนระหว่างอะตอมของกระดูกสันหลังของเปปไทด์
2. พันธะที่อ่อนแอระหว่างกลุ่มหน้าที่ของอนุมูลของกรดอะมิโน
3. พันธะระหว่างกรดอะมิโนกลุ่ม α-amino และ α-carboxyl
8. เลือกคำตอบที่ถูกต้อง. ทริปซิน:
ก. เอ็นไซม์โปรตีโอไลติก ข. ประกอบด้วยสองโดเมน
B. แป้งไฮโดรไลซ์
ง. ศูนย์ปฏิบัติการตั้งอยู่ระหว่างโดเมน ง. ประกอบด้วยสายโซ่โพลีเปปไทด์สองสาย
9. เลือกคำตอบที่ถูกต้อง. อะโทรพีน:
ก. สารสื่อประสาท
B. โครงสร้างอะนาล็อกของอะเซทิลโคลีน
B. โต้ตอบกับตัวรับ H-cholinergic
G. ช่วยเพิ่มการนำกระแสประสาทผ่าน cholinergic synapses
D. ตัวยับยั้งการแข่งขันของตัวรับ M-cholinergic
10. เลือกข้อความที่ถูกต้อง ในโปรตีน:
A. โครงสร้างหลักประกอบด้วยข้อมูลเกี่ยวกับโครงสร้างของไซต์ที่ใช้งานอยู่
B. ศูนย์แอคทีฟถูกสร้างขึ้นที่ระดับของโครงสร้างหลัก
B. โครงสร้างได้รับการแก้ไขอย่างเข้มงวดโดยพันธะโควาเลนต์
D. ไซต์ที่ใช้งานสามารถโต้ตอบกับกลุ่มของลิแกนด์ที่คล้ายคลึงกัน
เนื่องจากความสามารถในการกำหนดโครงสร้างของโปรตีน D. การเปลี่ยนแปลงสภาพแวดล้อมอาจส่งผลต่อความสัมพันธ์ของสารออกฤทธิ์
ศูนย์ลิแกนด์
1. 1-C, 2-D, 3-B.
3. A, B, C, D.
7. 1-B, 2-D, 3-A.
8. A, B, C, D.
ข้อกำหนดและแนวคิดพื้นฐาน
1. โปรตีน โพลีเปปไทด์ กรดอะมิโน
2. โครงสร้างโปรตีนระดับประถมศึกษา มัธยมศึกษา ระดับอุดมศึกษา
3. โครงสร้าง โครงสร้างโปรตีนพื้นเมือง
4. พันธะโควาเลนต์และอ่อนแอในโปรตีน
5. ความสามารถในการจัดรูปแบบ
6. ไซต์ที่ใช้งานโปรตีน
7. แกนด์
8. การพับโปรตีน
9. โครงสร้างแอนะล็อกของลิแกนด์
10. โดเมนโปรตีน
11. โปรตีนที่ง่ายและซับซ้อน
12. การเปลี่ยนสภาพโปรตีน, สารทำให้เสียสภาพ
13. การสร้างโปรตีนใหม่
แก้ปัญหา
"การจัดโครงสร้างโปรตีนและพื้นฐานของการทำงาน"
1. หน้าที่หลักของโปรตีน - เฮโมโกลบิน A (HbA) - คือการขนส่งออกซิเจนไปยังเนื้อเยื่อ ในประชากรมนุษย์ โปรตีนหลายรูปแบบนี้มีคุณสมบัติและหน้าที่เปลี่ยนแปลงไป ซึ่งเรียกว่าฮีโมโกลบินผิดปกติ ตัวอย่างเช่น ฮีโมโกลบิน S ที่พบในเม็ดเลือดแดงของผู้ป่วยที่มีโรคเม็ดเลือดรูปเคียว (HbS) พบว่ามีความสามารถในการละลายต่ำภายใต้สภาวะของความดันบางส่วนของออกซิเจนต่ำ (เช่นเดียวกับที่เกิดขึ้นในเลือดดำ) สิ่งนี้นำไปสู่การก่อตัวของมวลรวมของโปรตีนนี้ โปรตีนสูญเสียการทำงาน ตกตะกอน และเซลล์เม็ดเลือดแดงมีรูปร่างผิดปกติ (บางส่วนมีรูปร่างเป็นเคียว) และถูกทำลายเร็วกว่าปกติในม้าม เป็นผลให้เกิดโรคโลหิตจางเซลล์เคียว
ความแตกต่างเพียงอย่างเดียวในโครงสร้างหลักของ HvA พบได้ในบริเวณปลาย N ของสาย β ของเฮโมโกลบิน เปรียบเทียบบริเวณปลาย N ของสาย β และแสดงให้เห็นว่าการเปลี่ยนแปลงในโครงสร้างหลักของโปรตีนส่งผลต่อคุณสมบัติและหน้าที่ของโปรตีนอย่างไร
 สำหรับสิ่งนี้:
สำหรับสิ่งนี้:
ก) เขียนสูตรกรดอะมิโนโดยที่ HvA ต่างกันและเปรียบเทียบคุณสมบัติของกรดอะมิโนเหล่านี้ (ขั้ว ประจุ)
b) หาข้อสรุปเกี่ยวกับสาเหตุของการลดลงของความสามารถในการละลายและการละเมิดการขนส่งออกซิเจนในเนื้อเยื่อ
2. รูปแสดงแผนผังโครงสร้างของโปรตีนที่มีแกนด์ที่ยึดกับแกนกลาง (ศูนย์แอคทีฟ) อธิบายว่าเหตุใดจึงเลือกโปรตีนในการเลือกลิแกนด์ สำหรับสิ่งนี้:
ก) จำได้ว่าศูนย์กลางของโปรตีนคืออะไรและพิจารณาโครงสร้างของศูนย์กลางการทำงานของโปรตีนที่แสดงในรูป
b) เขียนสูตรของอนุมูลของกรดอะมิโนที่ประกอบขึ้นเป็นศูนย์ที่ใช้งานอยู่
c) วาดลิแกนด์ที่สามารถโต้ตอบอย่างเฉพาะเจาะจงกับไซต์ที่ทำงานของโปรตีน ระบุกลุ่มการทำงานที่สามารถสร้างพันธะกับอนุมูลของกรดอะมิโนที่ประกอบขึ้นเป็นศูนย์ที่ใช้งานอยู่
d) ระบุประเภทของพันธะที่เกิดขึ้นระหว่างลิแกนด์และอนุมูลของกรดอะมิโนของศูนย์แอคทีฟ
จ) อธิบายพื้นฐานสำหรับความจำเพาะของปฏิสัมพันธ์ของโปรตีนกับลิแกนด์
 3.
รูปแสดงตำแหน่งแอกทีฟไซต์ของโปรตีนและลิแกนด์หลายตัว
3.
รูปแสดงตำแหน่งแอกทีฟไซต์ของโปรตีนและลิแกนด์หลายตัว
พิจารณาว่าแกนด์ใดมีแนวโน้มที่จะโต้ตอบกับไซต์ที่ทำงานอยู่ของโปรตีนและทำไม
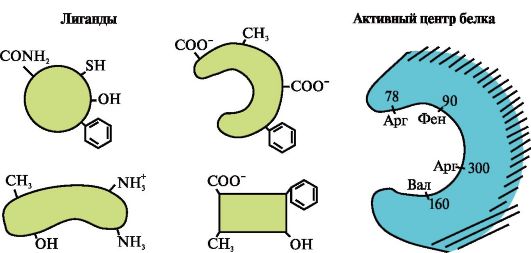 พันธะประเภทใดที่เกิดขึ้นระหว่างการก่อตัวของโปรตีนลิแกนด์คอมเพล็กซ์?
พันธะประเภทใดที่เกิดขึ้นระหว่างการก่อตัวของโปรตีนลิแกนด์คอมเพล็กซ์?
4. โครงสร้างแอนะล็อกของลิแกนด์โปรตีนธรรมชาติสามารถใช้เป็นยาในการเปลี่ยนแปลงกิจกรรมของโปรตีน
Acetylcholine เป็นตัวกลางในการส่งสัญญาณกระตุ้นในประสาทและกล้ามเนื้อประสาท เมื่อ acetylcholine ทำปฏิกิริยากับโปรตีน - ตัวรับของเยื่อหุ้มเซลล์ postsynaptic ของกล้ามเนื้อโครงร่าง ช่องไอออนจะเปิดขึ้นและเกิดการหดตัวของกล้ามเนื้อ Dithylin เป็นยาที่ใช้ในการผ่าตัดบางอย่างเพื่อผ่อนคลายกล้ามเนื้อ เนื่องจากจะไปขัดขวางการส่งสัญญาณของเส้นประสาทผ่านทางประสาทและกล้ามเนื้อ อธิบายกลไกการออกฤทธิ์ของไดไทลินเป็นยาคลายกล้ามเนื้อ สำหรับสิ่งนี้:
ก) เขียนสูตรของ acetylcholine และ dithyline และเปรียบเทียบโครงสร้างของพวกมัน
b) อธิบายกลไกของการกระทำที่ผ่อนคลายของ dithylin
5. ในบางโรค อุณหภูมิร่างกายของผู้ป่วยสูงขึ้น ซึ่งถือเป็นปฏิกิริยาป้องกันของร่างกาย อย่างไรก็ตาม อุณหภูมิสูงเป็นอันตรายต่อโปรตีนในร่างกาย อธิบายว่าเหตุใดที่อุณหภูมิสูงกว่า 40 °C การทำงานของโปรตีนจึงหยุดชะงักและเป็นอันตรายต่อชีวิตมนุษย์ เมื่อต้องการทำเช่นนี้ จำไว้ว่า:
1) โครงสร้างของโปรตีนและพันธะที่มีโครงสร้างอยู่ในรูปแบบดั้งเดิม
2) โครงสร้างและหน้าที่ของโปรตีนเปลี่ยนแปลงไปตามอุณหภูมิที่เพิ่มขึ้นอย่างไร;
3) สภาวะสมดุลคืออะไรและเหตุใดการรักษาสุขภาพของมนุษย์จึงมีความสำคัญ
ยูนิตโมดูลาร์ 2 OLIGOMERIC PROTEINS เป็นเป้าหมายสำหรับอิทธิพลด้านกฎระเบียบ โปรตีนหลากหลายโครงสร้างและหน้าที่การใช้งาน การแยกโปรตีนและวิธีการทำให้บริสุทธิ์
วัตถุประสงค์การเรียนรู้ เพื่อให้สามารถ:
1. ใช้ความรู้เกี่ยวกับคุณลักษณะของโครงสร้างและหน้าที่ของโปรตีน oligomeric เพื่อทำความเข้าใจกลไกการปรับตัวของการควบคุมการทำงานของพวกมัน
2. อธิบายบทบาทของพี่เลี้ยงในการสังเคราะห์และบำรุงรักษาโครงสร้างโปรตีนในเซลล์
3. เพื่ออธิบายความหลากหลายของการสำแดงชีวิตด้วยความหลากหลายของโครงสร้างและหน้าที่ของโปรตีนที่สังเคราะห์ขึ้นในร่างกาย
4. วิเคราะห์ความสัมพันธ์ระหว่างโครงสร้างของโปรตีนและหน้าที่ของโปรตีนโดยเปรียบเทียบเฮโมโกลบินที่เกี่ยวข้อง - ไมโอโกลบินและเฮโมโกลบิน ตลอดจนตัวแทนของโปรตีนห้าชั้นของตระกูลอิมมูโนโกลบูลิน
5. ใช้ความรู้เกี่ยวกับคุณสมบัติทางเคมีกายภาพของโปรตีนเพื่อเลือกวิธีการทำให้บริสุทธิ์จากโปรตีนและสิ่งเจือปนอื่นๆ
6. ตีความผลลัพธ์ขององค์ประกอบเชิงปริมาณและคุณภาพของโปรตีนในเลือดเพื่อยืนยันหรือชี้แจงการวินิจฉัยทางคลินิก
รู้:
1. คุณสมบัติของโครงสร้างของโปรตีน oligomeric และกลไกการปรับตัวของการควบคุมการทำงานของพวกมันในตัวอย่างของเฮโมโกลบิน
2. โครงสร้างและหน้าที่ของพี่เลี้ยงและความสำคัญในการรักษาโครงสร้างโปรตีนในเซลล์
3. หลักการจัดกลุ่มโปรตีนออกเป็นครอบครัวตามความคล้ายคลึงกันของโครงสร้างและหน้าที่ในตัวอย่างของอิมมูโนโกลบูลิน
4. วิธีการแยกโปรตีนตามคุณสมบัติของคุณสมบัติทางเคมีกายภาพ
5. อิเล็กโตรโฟรีซิสของพลาสมาเลือดเป็นวิธีการประเมินองค์ประกอบเชิงคุณภาพและเชิงปริมาณของโปรตีน
หัวข้อ 1.4. คุณสมบัติของโครงสร้างและการทำงานของโปรตีนโอลิโกเมอร์ในตัวอย่างของเฮโมโกลบิน
1. โปรตีนหลายชนิดประกอบด้วยสายโซ่โพลีเปปไทด์หลายสาย โปรตีนดังกล่าวเรียกว่า oligomeric,และแต่ละวงจร โปรโตเมอร์โปรโตเมอร์ในโปรตีนโอลิโกเมอร์เชื่อมต่อกันด้วยพันธะที่ไม่ใช่โควาเลนต์ที่อ่อนแอจำนวนมาก (ไม่ชอบน้ำ อิออน ไฮโดรเจน) ปฏิสัมพันธ์
protomers จะดำเนินการด้วย การเติมเต็มพื้นผิวสัมผัสของพวกเขา
จำนวนโปรโตเมอร์ในโปรตีนโอลิโกเมอร์อาจแตกต่างกันมาก: เฮโมโกลบินประกอบด้วย 4 โปรโตเมอร์ เอนไซม์แอสพาเทตอะมิโนทรานสเฟอเรส - 12 โปรโตเมอร์ และโปรตีนของไวรัสโมเสคยาสูบประกอบด้วยโปรโตเมอร์ 2120 ตัวที่เชื่อมต่อกันด้วยพันธะที่ไม่ใช่โควาเลนต์ ดังนั้นโปรตีนโอลิโกเมอร์จึงสามารถมีน้ำหนักโมเลกุลสูงมาก
ปฏิสัมพันธ์ของโปรโตเมอร์หนึ่งกับคนอื่นถือได้ว่าเป็นกรณีพิเศษของปฏิสัมพันธ์ของโปรตีนกับลิแกนด์เนื่องจากโปรโตเมอร์แต่ละตัวทำหน้าที่เป็นลิแกนด์สำหรับโปรโตเมอร์อื่น จำนวนและวิธีการเชื่อมต่อโปรโตเมอร์ในโปรตีนเรียกว่า โครงสร้างโปรตีนสี่ส่วน
โปรตีนสามารถมีโปรโตเมอร์ที่มีโครงสร้างเหมือนกันหรือต่างกัน ตัวอย่างเช่น โฮโมไดเมอร์คือโปรตีนที่มีโปรโตเมอร์ที่เหมือนกันสองตัว และเฮเทอโรไดเมอร์คือโปรตีนที่มีโปรโตเมอร์ที่ต่างกันสองตัว
หากโปรตีนมีโปรโตเมอร์ต่างกัน ศูนย์การจับกับลิแกนด์ที่แตกต่างกันซึ่งมีโครงสร้างต่างกันก็สามารถก่อตัวขึ้นได้ เมื่อลิแกนด์จับกับศูนย์กลางที่ทำงานอยู่ หน้าที่ของโปรตีนนี้จะปรากฏออกมา ศูนย์ที่อยู่บนโปรโตเมอร์อื่นเรียกว่าอัลโลสเตอริก (นอกเหนือจากแอคทีฟ) การติดต่อ ลิแกนด์อัลโลสเตอริกหรือเอฟเฟกเตอร์มันทำหน้าที่กำกับดูแล (รูปที่ 1.18) การทำงานร่วมกันของศูนย์ allosteric กับเอฟเฟกต์ทำให้เกิดการเปลี่ยนแปลงเชิงโครงสร้างในโครงสร้างของโปรตีน oligomeric ทั้งหมดเนื่องจากความสามารถในการกำหนดรูปแบบ สิ่งนี้ส่งผลต่อสัมพรรคภาพของไซต์แอคทีฟสำหรับลิแกนด์จำเพาะและควบคุมการทำงานของโปรตีนนั้น การเปลี่ยนแปลงโครงสร้างและหน้าที่ของโปรโตเมอร์ทั้งหมดระหว่างอันตรกิริยาของโปรตีนโอลิโกเมอร์กับลิแกนด์อย่างน้อยหนึ่งตัวเรียกว่าการเปลี่ยนแปลงเชิงโครงสร้างแบบร่วมมือ เอฟเฟคเตอร์ที่เสริมการทำงานของโปรตีนเรียกว่า ตัวกระตุ้นและเอฟเฟกต์ที่กดดันการทำงานของมัน - สารยับยั้ง
ดังนั้นในโปรตีน oligomeric เช่นเดียวกับโปรตีนที่มีโครงสร้างโดเมน คุณสมบัติใหม่จะปรากฏขึ้นเมื่อเปรียบเทียบกับโปรตีนโมโนเมอร์ - ความสามารถในการควบคุมการทำงานแบบ allosterically (การควบคุมโดยแนบลิแกนด์ที่แตกต่างกันกับโปรตีน) สิ่งนี้สามารถเห็นได้โดยการเปรียบเทียบโครงสร้างและหน้าที่ของโปรตีนที่ซับซ้อนสองชนิดที่เกี่ยวข้องกันอย่างใกล้ชิดคือไมโอโกลบินและเฮโมโกลบิน
 ข้าว. 1.18. แผนภาพโครงสร้างของโปรตีนไดเมอริก
ข้าว. 1.18. แผนภาพโครงสร้างของโปรตีนไดเมอริก
2. การก่อตัวของโครงสร้างเชิงพื้นที่และการทำงานของ myoglobin
Myoglobin (Mb) เป็นโปรตีนที่พบในกล้ามเนื้อสีแดง หน้าที่หลักคือการสร้าง O 2 สำรองที่จำเป็นสำหรับการทำงานของกล้ามเนื้อที่รุนแรง MB เป็นโปรตีนเชิงซ้อนที่มีส่วนโปรตีน - apoMB และส่วนที่ไม่ใช่โปรตีน - heme โครงสร้างหลักของ apoMB กำหนดโครงสร้างทรงกลมขนาดกะทัดรัดและโครงสร้างของศูนย์แอ็คทีฟซึ่งแนบส่วนที่ไม่ใช่โปรตีนของ myoglobin, heme ออกซิเจนจากเลือดไปยังกล้ามเนื้อจับกับ Fe + 2 heme ในองค์ประกอบของ myoglobin MB เป็นโปรตีนโมโนเมอร์ที่มีความสัมพันธ์กับ O 2 สูงมาก ดังนั้น myoglobin จะปล่อยออกซิเจนเฉพาะในระหว่างการทำงานของกล้ามเนื้อที่รุนแรงเท่านั้น เมื่อความดันบางส่วนของ O 2 ลดลงอย่างรวดเร็ว
การก่อตัวของรูปแบบ MB.ในกล้ามเนื้อสีแดง บนไรโบโซมระหว่างการแปล การสังเคราะห์โครงสร้างหลักของ MB ซึ่งแสดงโดยลำดับเฉพาะของ 153 กรดอะมิโนตกค้างเกิดขึ้น โครงสร้างทุติยภูมิของ Mv ประกอบด้วย α-helices แปดตัว เรียกว่าตัวอักษรละตินจาก A ถึง H ซึ่งจะมีส่วนที่ไม่เป็นเกลียว โครงสร้างระดับตติยภูมิของ Mv มีรูปทรงกลมขนาดกะทัดรัด ซึ่งอยู่ระหว่าง F และ E α-helices มีจุดศูนย์กลางที่แอคทีฟ (รูปที่ 1.19)
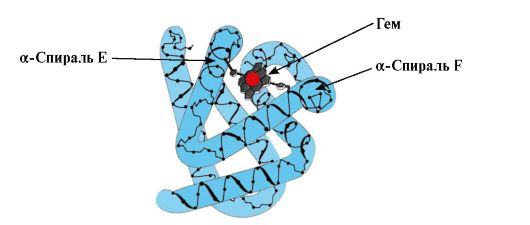 ข้าว. 1.19. โครงสร้างของไมโอโกลบิน
ข้าว. 1.19. โครงสร้างของไมโอโกลบิน
3. คุณสมบัติของโครงสร้างและการทำงานของศูนย์แอคทีฟ MVศูนย์กลางแอคทีฟของ Mv ส่วนใหญ่เกิดจากอนุมูลของกรดอะมิโนที่ไม่ชอบน้ำซึ่งอยู่ห่างไกลจากกันในโครงสร้างปฐมภูมิ (เช่น Tri 3 9 และเพ็ญ 138) ลิแกนด์ที่ละลายได้ไม่ดีในน้ำ heme และ O 2 ติดอยู่ที่ศูนย์แอคทีฟ Heme เป็นลิแกนด์ apoMv เฉพาะ (รูปที่ 1.20) ซึ่งอิงจากวงแหวนไพร์โรลสี่วงที่เชื่อมต่อด้วยสะพานเมธินิล ตรงกลางมีอะตอม Fe+ 2 เชื่อมต่อกับอะตอมไนโตรเจนของวงแหวนไพร์โรลด้วยพันธะประสาน 4 พันธะ นอกจากอนุมูลที่ไม่ชอบน้ำของกรดอะมิโนแล้ว ศูนย์กลางของ Mv ยังประกอบด้วยสารตกค้างของกรดอะมิโนสองชนิดที่มีอนุมูลที่ชอบน้ำ - Gis E7(GIS 64) และ Gis F 8(พระองค์ 93) (รูปที่ 1.21).
 ข้าว. 1.20. โครงสร้างของ heme - ส่วนที่ไม่ใช่โปรตีนของ myoglobin และ hemoglobin
ข้าว. 1.20. โครงสร้างของ heme - ส่วนที่ไม่ใช่โปรตีนของ myoglobin และ hemoglobin
 ข้าว. 1.21. ตำแหน่งของ heme และ O 2 ในบริเวณแอกทีฟไซต์ของ apomyoglobin และ hemoglobin protomers
ข้าว. 1.21. ตำแหน่งของ heme และ O 2 ในบริเวณแอกทีฟไซต์ของ apomyoglobin และ hemoglobin protomers
Heme ถูกพันธะโควาเลนต์กับ F 8 ของเขาผ่านอะตอมของเหล็ก O 2 ยึดติดกับเหล็กที่อีกด้านหนึ่งของระนาบ heme E 7 ของเขาจำเป็นสำหรับการวางแนวที่ถูกต้องของ O 2 และอำนวยความสะดวกในการเพิ่มออกซิเจนให้กับ Fe + 2 heme
Gis F 8สร้างพันธะประสานงานกับ Fe+ 2 และแก้ไข heme อย่างแน่นหนาในศูนย์ที่ใช้งานอยู่ Gis E7จำเป็นสำหรับการวางแนวที่ถูกต้องในศูนย์กลางของแกนด์อื่น - O 2 ในระหว่างการโต้ตอบกับ Fe + 2 heme สภาวะแวดล้อมจุลภาคของ heme สร้างสภาวะสำหรับการยึดเกาะที่แข็งแรงแต่สามารถย้อนกลับของ O 2 กับ Fe + 2 และป้องกันไม่ให้น้ำเข้าสู่ศูนย์แอคทีฟที่ไม่ชอบน้ำ ซึ่งสามารถนำไปสู่การออกซิเดชันของ Fe + 3 ได้
โครงสร้างโมโนเมอร์ของ MB และศูนย์กลางที่แอคทีฟกำหนดความสัมพันธ์สูงของโปรตีนสำหรับ O 2
4. โครงสร้าง Oligomeric ของ Hb และการควบคุมความสัมพันธ์ของ Hb สำหรับ O 2 โดยลิแกนด์ ฮีโมโกลบินของมนุษย์- ตระกูลของโปรตีน เช่นเดียวกับ myoglobin ที่เกี่ยวข้องกับโปรตีนเชิงซ้อน (hemoproteins) พวกมันมีโครงสร้างเตตระเมอร์และประกอบด้วยสาย α สองสาย แต่โครงสร้างของสายโพลีเปปไทด์อีกสองสายต่างกัน (2α-, 2x-chains) โครงสร้างของสายพอลิเปปไทด์ที่สองกำหนดคุณลักษณะของการทำงานของรูปแบบเหล่านี้ของ Hb ประมาณ 98% ของฮีโมโกลบินในเม็ดเลือดแดงของผู้ใหญ่คือ เฮโมโกลบิน A(2α-, 2p-เชน)
ในระหว่างการพัฒนาของทารกในครรภ์มีฮีโมโกลบินสองประเภทหลัก: HB .ตัวอ่อน(2α, 2ε) ซึ่งพบได้ในระยะแรกของการพัฒนาของทารกในครรภ์และ เฮโมโกลบิน F (ทารกในครรภ์)- (2α, 2γ) ซึ่งแทนที่ฮีโมโกลบินของทารกในครรภ์ในระยะแรกในเดือนที่หกของการพัฒนาของทารกในครรภ์และจะถูกแทนที่ด้วย Hb A หลังคลอดเท่านั้น
Hv A เป็นโปรตีนที่เกี่ยวข้องกับ myoglobin (Mv) ที่พบในเม็ดเลือดแดงของผู้ใหญ่ โครงสร้างของโปรโตเมอร์แต่ละตัวนั้นคล้ายกับของไมโอโกลบิน โครงสร้างทุติยภูมิและตติยภูมิของ myoglobin และ hemoglobin protomers มีความคล้ายคลึงกันมาก แม้ว่าจะมีกรดอะมิโนเหลืออยู่เพียง 24 ชนิดเท่านั้นที่เหมือนกันในโครงสร้างหลักของสายโพลีเปปไทด์ (โครงสร้างรองของโปรโตเมอร์เฮโมโกลบิน เช่น myoglobin มี α-helices อยู่แปดตัว แสดงด้วยตัวอักษรละตินจาก A ถึง H และโครงสร้างตติยภูมิมีรูปทรงกลมขนาดกะทัดรัด) แต่แตกต่างจาก myoglobin ตรงที่ เฮโมโกลบินมีโครงสร้างโอลิโกเมอร์ ประกอบด้วยสายโซ่โพลีเปปไทด์สี่สายที่เชื่อมต่อกันด้วยพันธะที่ไม่ใช่โควาเลนต์ (รูปที่ 1.22)
โปรโตเมอร์ Hb แต่ละตัวมีความเกี่ยวข้องกับส่วนที่ไม่ใช่โปรตีน - ฮีมและโปรโตเมอร์ที่อยู่ใกล้เคียง การเชื่อมต่อของส่วนโปรตีนของ Hb กับ heme นั้นคล้ายกับของ myoglobin: ในศูนย์กลางของโปรตีนส่วนที่ไม่ชอบน้ำของ heme นั้นล้อมรอบด้วยอนุมูลของกรดอะมิโนที่ไม่ชอบน้ำ ยกเว้น His F 8 และ His E 7 ซึ่งตั้งอยู่ทั้งสองด้านของระนาบ heme และมีบทบาทคล้ายกันในการทำงานของโปรตีนและการจับกับออกซิเจน (ดูโครงสร้างของ myoglobin)
 ข้าว. 1.22. โครงสร้าง Oligomeric ของเฮโมโกลบิน
ข้าว. 1.22. โครงสร้าง Oligomeric ของเฮโมโกลบิน
นอกจากนี้, Gis E7ทำหน้าที่สำคัญ บทบาทเพิ่มเติมในการทำงานของ NV ฟรี heme มีความสัมพันธ์ใกล้ชิดกับ CO มากกว่า O 2 ถึง 25,000 เท่า CO ก่อตัวขึ้นในปริมาณเล็กน้อยในร่างกาย และเนื่องจากมีความเกี่ยวข้องกับ heme สูง จึงอาจขัดขวางการลำเลียง O 2 ที่จำเป็นสำหรับชีวิตของเซลล์ อย่างไรก็ตาม ในองค์ประกอบของเฮโมโกลบิน ความสัมพันธ์ของฮีมต่อคาร์บอนมอนอกไซด์นั้นสูงกว่าค่าสัมพรรคภาพของ O 2 เพียง 200 เท่าเนื่องจากการมีอยู่ของ E 7 ในศูนย์กลางที่แอคทีฟของเขา ส่วนที่เหลือของกรดอะมิโนนี้จะสร้างสภาวะที่เหมาะสมที่สุดสำหรับการจับตัวของ heme กับ O2 และทำให้ปฏิกิริยาของ heme กับ CO อ่อนแอลง
5. หน้าที่หลักของ Hb คือการขนส่ง O 2 จากปอดไปยังเนื้อเยื่อโครงสร้าง oligomeric ของเฮโมโกลบินไม่เหมือนกับโมโนเมอร์ที่มีความสัมพันธ์กับ O 2 และทำหน้าที่เก็บออกซิเจนในกล้ามเนื้อสีแดง:
1) ความอิ่มตัวอย่างรวดเร็วของ Hb กับออกซิเจนในปอด
2) ความสามารถของ Hb ในการปลดปล่อยออกซิเจนในเนื้อเยื่อที่ความดันบางส่วนที่ค่อนข้างสูงของ O 2 (20-40 mm Hg)
3) ความเป็นไปได้ในการควบคุมความสัมพันธ์ของ Hb กับ O 2 .
6. การเปลี่ยนแปลงร่วมกันในโครงสร้างของโปรโตเมอร์ของเฮโมโกลบินเร่งการผูกมัดของ O 2 ในปอดและการกลับสู่เนื้อเยื่อ ในปอด ความดันบางส่วนที่สูงของ O2 ส่งเสริมการผูกมัดกับ Hb ในบริเวณที่ทำงานของโปรโตเมอร์สี่ตัว (2αและ2β) จุดศูนย์กลางที่แอคทีฟของโปรโตเมอร์แต่ละตัว เช่นเดียวกับใน myoglobin นั้นตั้งอยู่ระหว่าง α-helices (F และ E) สองตัวในกระเป๋าที่ไม่ชอบน้ำ ประกอบด้วยส่วนที่ไม่ใช่โปรตีน - heme ติดกับส่วนโปรตีนโดยปฏิกิริยาที่ไม่ชอบน้ำที่อ่อนแอจำนวนมาก และพันธะที่แน่นหนาระหว่าง Fe 2 + heme กับ His F 8 (ดูรูปที่ 1.21)
ใน deoxyhemoglobin เนื่องจากการเชื่อมต่อกับ His F 8 อะตอม Fe 2 + จะยื่นออกมาจากระนาบ heme ไปทางฮิสทิดีน การผูกมัดของ O 2 กับ Fe 2 + เกิดขึ้นที่อีกด้านหนึ่งของ heme ในภูมิภาค His E 7 ด้วยความช่วยเหลือของพันธะประสานงานอิสระเดียว E 7 ของเขาให้สภาวะที่เหมาะสมที่สุดสำหรับการผูก O 2 ด้วยเหล็กเฮม
การเพิ่ม O 2 เข้ากับอะตอม Fe +2 ของโปรโตเมอร์หนึ่งตัวทำให้เคลื่อนเข้าสู่ระนาบ heme และด้านหลังมีฮิสทิดีนตกค้างที่เกี่ยวข้อง
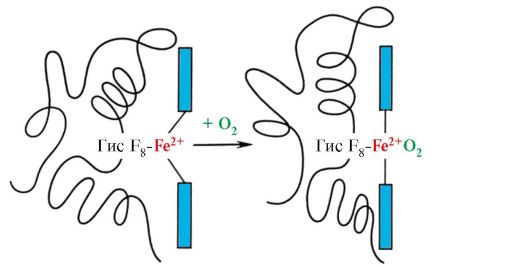 ข้าว. 1.23. การเปลี่ยนแปลงรูปร่างของโปรโตเมอร์ของฮีโมโกลบินเมื่อรวมกับ O 2
ข้าว. 1.23. การเปลี่ยนแปลงรูปร่างของโปรโตเมอร์ของฮีโมโกลบินเมื่อรวมกับ O 2
สิ่งนี้นำไปสู่การเปลี่ยนแปลงในโครงสร้างของสายโพลีเปปไทด์ทั้งหมดเนื่องจากความสามารถในการกำหนดรูปแบบได้ การเปลี่ยนโครงสร้างของสายโซ่อื่นๆ ช่วยให้เกิดปฏิสัมพันธ์กับโมเลกุล O 2 ถัดไปได้ง่ายขึ้น
โมเลกุล O 2 ที่สี่ยึดติดกับเฮโมโกลบินได้ง่ายกว่าโมเลกุลแรก 300 เท่า (รูปที่ 1.24)
 ข้าว. 1.24. การเปลี่ยนแปลงร่วมกันในโครงสร้างของโปรโตเมอร์เฮโมโกลบินในระหว่างการมีปฏิสัมพันธ์กับ O 2
ข้าว. 1.24. การเปลี่ยนแปลงร่วมกันในโครงสร้างของโปรโตเมอร์เฮโมโกลบินในระหว่างการมีปฏิสัมพันธ์กับ O 2
ในเนื้อเยื่อ แต่ละโมเลกุลของ O 2 ที่ตามมาจะถูกตัดออกได้ง่ายกว่าโมเลกุลก่อนหน้า นอกจากนี้ เนื่องจากการเปลี่ยนแปลงร่วมกันในโครงสร้างของโปรโตเมอร์
7. CO 2 และ H + เกิดขึ้นระหว่าง catabolism ของสารอินทรีย์ ลดความสัมพันธ์ของฮีโมโกลบินสำหรับ O 2 ตามสัดส่วนของความเข้มข้น พลังงานที่จำเป็นสำหรับการทำงานของเซลล์ส่วนใหญ่ผลิตขึ้นในไมโตคอนเดรียในระหว่างการออกซิเดชันของสารอินทรีย์โดยใช้ O 2 ที่ส่งมาจากปอดโดยเฮโมโกลบิน เป็นผลมาจากการเกิดออกซิเดชันของสารอินทรีย์ทำให้เกิดผลิตภัณฑ์ขั้นสุดท้ายของการสลายตัว: CO 2 และ K 2 O ปริมาณซึ่งเป็นสัดส่วนกับความเข้มของกระบวนการออกซิเดชันอย่างต่อเนื่อง
CO 2 แพร่กระจายจากเซลล์ไปสู่กระแสเลือดและเข้าสู่เม็ดเลือดแดงซึ่งภายใต้การกระทำของเอนไซม์ carbanhydrase จะเปลี่ยนเป็นกรดคาร์บอนิก กรดอ่อนนี้จะแยกตัวออกเป็นโปรตอนและไอออนไบคาร์บอเนต
H+ สามารถเข้าร่วมอนุมูล GIS ได้ 14 6 ใน α- และ β-chains ของเฮโมโกลบินเช่น ในพื้นที่ห่างไกลจากเฮม โปรตอนของเฮโมโกลบินลดความสัมพันธ์กับ O 2 ส่งเสริมการกำจัด O 2 ออกจาก oxyHb การก่อตัวของ deoxyHb และเพิ่มปริมาณออกซิเจนไปยังเนื้อเยื่อตามสัดส่วนของจำนวนโปรตอนที่เกิดขึ้น (รูปที่ 1.25)
การเพิ่มขึ้นของปริมาณออกซิเจนที่ปล่อยออกมาขึ้นอยู่กับการเพิ่มขึ้นของความเข้มข้นของ H + ในเม็ดเลือดแดงเรียกว่าเอฟเฟกต์ Bohr (หลังจากนักสรีรวิทยาชาวเดนมาร์ก Christian Bohr ผู้ค้นพบผลกระทบนี้เป็นครั้งแรก)
ในปอด ความดันบางส่วนของออกซิเจนสูงจะส่งเสริมการจับกับ deoxyHb ซึ่งจะช่วยลดความสัมพันธ์ของโปรตีนสำหรับ H+ โปรตอนที่ถูกปล่อยออกมาภายใต้การกระทำของ carbanhydrase ทำปฏิกิริยากับไบคาร์บอเนตเพื่อสร้าง CO 2 และ H 2 O

 ข้าว. 1.25. การพึ่งพาความสัมพันธ์ของ Hb กับ O 2 กับความเข้มข้นของ CO 2 และโปรตอน (ผลกระทบจากบอร์):
ข้าว. 1.25. การพึ่งพาความสัมพันธ์ของ Hb กับ O 2 กับความเข้มข้นของ CO 2 และโปรตอน (ผลกระทบจากบอร์):
แต่- อิทธิพลของความเข้มข้นของ CO 2 และ H+ ต่อการปล่อย O 2 จากคอมเพล็กซ์ด้วย Hb (เอฟเฟกต์บอร์) บี- การเติมออกซิเจนของดีออกซีเฮโมโกลบินในปอด การก่อตัวและการปล่อย CO 2
CO 2 ที่เป็นผลลัพธ์จะเข้าสู่ช่องว่างของถุงลมและถูกขับออกด้วยอากาศที่หายใจออก ดังนั้นปริมาณออกซิเจนที่ปล่อยออกมาจากเฮโมโกลบินในเนื้อเยื่อจึงถูกควบคุมโดยผลิตภัณฑ์ของแคแทบอลิซึมของสารอินทรีย์: ยิ่งการสลายตัวของสารรุนแรงขึ้นเช่นในระหว่างการออกแรงทางกายภาพความเข้มข้นของ CO 2 และ H + ยิ่งสูงขึ้น ออกซิเจนที่เนื้อเยื่อได้รับเนื่องจากความสัมพันธ์ระหว่าง H ถึง O 2 ลดลง
8. ระเบียบ Allosteric ของความสัมพันธ์ของ Hb สำหรับ O 2 โดยลิแกนด์ - 2,3-bisphosphoglycerateในเม็ดเลือดแดง ลิแกนด์ allosteric ของเฮโมโกลบิน 2,3-bisphosphoglycerate (2,3-BPG) ถูกสังเคราะห์จากผลิตภัณฑ์ของการเกิดออกซิเดชันของกลูโคส - 1,3-bisphosphoglycerate ภายใต้สภาวะปกติ ความเข้มข้นของ 2,3-BPG จะสูงและเทียบได้กับความเข้มข้นของ Hb 2,3-BPG มีประจุลบที่แข็งแกร่งที่ -5
 Bisphosphoglycerate ในเนื้อเยื่อเส้นเลือดฝอย โดยการจับกับ deoxyhemoglobin จะเพิ่มปริมาณออกซิเจนในเนื้อเยื่อ ลดความสัมพันธ์ของ Hb ต่อ O 2
Bisphosphoglycerate ในเนื้อเยื่อเส้นเลือดฝอย โดยการจับกับ deoxyhemoglobin จะเพิ่มปริมาณออกซิเจนในเนื้อเยื่อ ลดความสัมพันธ์ของ Hb ต่อ O 2
มีโพรงอยู่ตรงกลางของโมเลกุลเฮโมโกลบินเตตราเมอร์ มันถูกสร้างขึ้นโดยส่วนที่เหลือของกรดอะมิโนของโปรโตเมอร์ทั้งสี่ (ดูรูปที่ 1.22) ในเส้นเลือดฝอยของเนื้อเยื่อ โปรตอนของ Hb (ผลกระทบจากบอร์) จะทำลายพันธะระหว่างธาตุเหล็ก heme และ O 2 ในโมเลกุล
deoxyhemoglobin เมื่อเทียบกับ oxyhemoglobin พันธะไอออนิกเพิ่มเติมจะปรากฏขึ้นที่เชื่อมต่อโปรโตเมอร์อันเป็นผลมาจากขนาดของโพรงกลางเพิ่มขึ้นเมื่อเทียบกับ oxyhemoglobin ช่องกลางคือบริเวณที่ยึด 2,3-BPG กับเฮโมโกลบิน เนื่องจากความแตกต่างของขนาดของช่องกลาง 2,3-BPG สามารถยึดติดกับ deoxyhemoglobin เท่านั้น
2,3-BPG ทำปฏิกิริยากับเฮโมโกลบินในพื้นที่ห่างไกลจากไซต์แอคทีฟของโปรตีนและเป็นของ allosteric(ระเบียบข้อบังคับ) ลิแกนด์และช่องกลาง Hb is ศูนย์อัลลอสเตอร์ 2,3-BPG มีประจุลบรุนแรงและมีปฏิสัมพันธ์กับกลุ่มที่มีประจุบวกห้ากลุ่มของสาย Hb β 2 กลุ่ม: กลุ่ม α-amino ของปลาย N-term Val และ Lys 82 Gis 143 เรดิคัล (รูปที่ 1.26)
 ข้าว. 1.26. BPG ในช่องกลางของ deoxyhemoglobin
ข้าว. 1.26. BPG ในช่องกลางของ deoxyhemoglobin
BPG จับกับกลุ่มที่มีประจุบวกสามกลุ่มในแต่ละ β-strand
ในเนื้อเยื่อเส้นเลือดฝอย ผลที่ได้คือดีออกซีเฮโมโกลบินมีปฏิสัมพันธ์กับ 2,3-BPG และพันธะไอออนิกเกิดขึ้นระหว่างอนุมูลที่มีประจุบวกของสาย β และลิแกนด์ที่มีประจุลบ ซึ่งเปลี่ยนรูปแบบโปรตีนและลดความสัมพันธ์ของ Hb สำหรับ O 2 ความสัมพันธ์ที่ลดลงของ Hb สำหรับ O 2 มีส่วนทำให้การปลดปล่อย O 2 เข้าสู่เนื้อเยื่อมีประสิทธิภาพมากขึ้น
ในปอด ที่ความดันบางส่วนสูง ออกซิเจนจะทำปฏิกิริยากับ Hb รวมเข้ากับเหล็กฮีม ในกรณีนี้ โครงสร้างของโปรตีนจะเปลี่ยนไป ช่องกลางลดลง และ 2,3-BPG ถูกแทนที่จากศูนย์กลางอัลโลสเตอริก
ดังนั้นโปรตีนโอลิโกเมอร์จึงมีคุณสมบัติใหม่เมื่อเปรียบเทียบกับโปรตีนโมโนเมอร์ สิ่งที่แนบมาของลิแกนด์ที่ไซต์
อยู่ห่างจากกันเชิงพื้นที่ (อัลลอสเตอริก) ซึ่งสามารถทำให้เกิดการเปลี่ยนแปลงเชิงโครงสร้างในโมเลกุลโปรตีนทั้งหมด เนื่องจากปฏิสัมพันธ์กับลิแกนด์ควบคุม การเปลี่ยนแปลงรูปแบบและการทำงานของโมเลกุลโปรตีนจึงปรับให้เข้ากับการเปลี่ยนแปลงของสิ่งแวดล้อม
หัวข้อ 1.5. การบำรุงรักษาโครงสร้างโปรตีนโดยกำเนิดภายใต้สภาวะของเซลล์
ในเซลล์ ในระหว่างการสังเคราะห์สายโซ่โพลีเปปไทด์ การขนส่งผ่านเยื่อหุ้มเซลล์ไปยังส่วนที่เกี่ยวข้องของเซลล์ ในกระบวนการพับ (การก่อตัวของโครงสร้างดั้งเดิม) และระหว่างการประกอบโปรตีนโอลิโกเมอร์ตลอดจนระหว่างการทำงาน ระดับกลาง , โครงสร้างที่มีแนวโน้มการรวมตัว, ที่ไม่เสถียรเกิดขึ้นในโครงสร้างโปรตีน อนุมูลที่ไม่ชอบน้ำ ซึ่งมักจะซ่อนอยู่ภายในโมเลกุลโปรตีนในรูปแบบดั้งเดิม ปรากฏบนพื้นผิวในรูปแบบที่ไม่เสถียรและมีแนวโน้มที่จะรวมกับกลุ่มของโปรตีนอื่นๆ ที่ละลายได้ไม่ดีในน้ำในทำนองเดียวกัน ในเซลล์ของสิ่งมีชีวิตที่รู้จักทั้งหมด มีการค้นพบโปรตีนพิเศษที่ให้การพับของโปรตีนในเซลล์อย่างเหมาะสม รักษาเสถียรภาพของโครงสร้างดั้งเดิมระหว่างการทำงาน และที่สำคัญที่สุดคือ รักษาโครงสร้างและหน้าที่ของโปรตีนภายในเซลล์ในกรณีที่เกิดภาวะธำรงดุลยภาพผิดปกติ โปรตีนเหล่านี้เรียกว่า "พี่เลี้ยง"ซึ่งหมายถึง "พี่เลี้ยง" ในภาษาฝรั่งเศส
1. พี่เลี้ยงระดับโมเลกุลและบทบาทในการป้องกันการเสื่อมสภาพของโปรตีน
พี่เลี้ยง (III) จำแนกตามมวลของหน่วยย่อย พี่เลี้ยงที่มีน้ำหนักโมเลกุลสูงมีมวล 60 ถึง 110 kD ในหมู่พวกเขามีสามชั้นเรียนที่ได้รับการศึกษามากที่สุด: Sh-60, Sh-70 และ Sh-90 แต่ละชั้นประกอบด้วยตระกูลของโปรตีนที่เกี่ยวข้อง ดังนั้น Sh-70 จึงมีโปรตีนที่มีน้ำหนักโมเลกุล 66 ถึง 78 kD พี่เลี้ยงที่มีน้ำหนักโมเลกุลต่ำมีน้ำหนักโมเลกุล 40 ถึง 15 kD
ในบรรดาพี่เลี้ยงมี รัฐธรรมนูญโปรตีนที่มีการสังเคราะห์พื้นฐานสูงไม่ขึ้นอยู่กับผลกระทบต่อเซลล์ของร่างกายและ เหนี่ยวนำให้เกิด,การสังเคราะห์ซึ่งภายใต้สภาวะปกตินั้นอ่อนแอ แต่เพิ่มขึ้นอย่างรวดเร็วภายใต้อิทธิพลที่ตึงเครียด พี่เลี้ยงที่เหนี่ยวนำได้เรียกอีกอย่างว่า "โปรตีนช็อตจากความร้อน" เนื่องจากถูกค้นพบครั้งแรกในเซลล์ที่สัมผัสกับ อุณหภูมิสูง. ในเซลล์ เนื่องจากโปรตีนมีความเข้มข้นสูง การงอกใหม่ตามธรรมชาติของโปรตีนที่เสียสภาพบางส่วนจึงเป็นเรื่องยาก Sh-70 สามารถป้องกันกระบวนการทำให้เสียสภาพที่เริ่มขึ้นและช่วยฟื้นฟูโครงสร้างโปรตีนตามธรรมชาติ โมเลกุลพี่เลี้ยง-70- คลาสโปรตีนที่มีการอนุรักษ์อย่างสูงที่พบในทุกส่วนของเซลล์: ไซโตพลาสซึม, นิวเคลียส, เอนโดพลาสมิกเรติคูลัม, ไมโทคอนเดรีย ที่ปลายคาร์บอกซิลของสายโซ่พอลิเปปไทด์เพียงสายเดียวของ Sh-70 มีบริเวณที่เป็นร่องที่สามารถโต้ตอบกับเปปไทด์ที่มีความยาวได้
จากสารตกค้างของกรดอะมิโน 7 ถึง 9 ตัวที่อุดมไปด้วยอนุมูลอิสระ ตำแหน่งดังกล่าวในโปรตีนทรงกลมเกิดขึ้นทุกๆ 16 กรดอะมิโนโดยประมาณ Sh-70 สามารถปกป้องโปรตีนจากการยับยั้งความร้อนและฟื้นฟูโครงสร้างและกิจกรรมของโปรตีนที่เสียสภาพบางส่วน
2. บทบาทของพี่เลี้ยงในการพับโปรตีนในระหว่างการสังเคราะห์โปรตีนบนไรโบโซม บริเวณปลาย N ของโพลีเปปไทด์จะถูกสังเคราะห์ก่อนบริเวณปลาย C ลำดับกรดอะมิโนที่สมบูรณ์ของโปรตีนจำเป็นต่อการก่อรูปโครงสร้างโดยกำเนิด ในกระบวนการสังเคราะห์โปรตีน พี่เลี้ยง-70 เนื่องจากโครงสร้างของจุดศูนย์กลางที่แอคทีฟของพวกมัน จึงสามารถปิดบริเวณที่มีแนวโน้มการรวมตัวของโพลีเปปไทด์ที่อุดมไปด้วยอนุมูลของกรดอะมิโนที่ไม่ชอบน้ำได้จนกว่าการสังเคราะห์จะเสร็จสิ้น (รูปที่ 1.27, A)
 ข้าว. 1.27. การมีส่วนร่วมของพี่เลี้ยงในการพับโปรตีน
ข้าว. 1.27. การมีส่วนร่วมของพี่เลี้ยงในการพับโปรตีน
เอ - การมีส่วนร่วมของพี่เลี้ยง-70 ในการป้องกันปฏิกิริยาที่ไม่ชอบน้ำระหว่างไซต์ของโพลีเปปไทด์สังเคราะห์ B - การก่อตัวของรูปแบบโปรตีนพื้นเมืองในคอมเพล็กซ์พี่เลี้ยง
โปรตีนที่มีน้ำหนักโมเลกุลสูงจำนวนมากที่มีโครงสร้างที่ซับซ้อน เช่น โครงสร้างโดเมน พับในพื้นที่พิเศษที่สร้างโดย W-60 Sh-60ทำหน้าที่เป็นคอมเพล็กซ์ oligomeric ซึ่งประกอบด้วย 14 หน่วยย่อย พวกเขาสร้างวงแหวนกลวงสองวงซึ่งแต่ละอันประกอบด้วยเจ็ดหน่วยย่อยวงแหวนเหล่านี้เชื่อมต่อกัน แต่ละหน่วยย่อยของ III-60 ประกอบด้วยสามโดเมน: ปลาย (ปลาย) อุดมด้วยอนุมูลที่ไม่ชอบน้ำซึ่งหันหน้าไปทางโพรงของวงแหวน ระดับกลางและเส้นศูนย์สูตร (รูปที่ 1.28)
 ข้าว. 1.28. โครงสร้างของ chaperonin complex ประกอบด้วย 14 Sh-60
ข้าว. 1.28. โครงสร้างของ chaperonin complex ประกอบด้วย 14 Sh-60
เอ - มุมมองด้านข้าง; B - มุมมองด้านบน
โปรตีนสังเคราะห์ที่มีองค์ประกอบพื้นผิวที่มีลักษณะเฉพาะของโมเลกุลที่กางออกโดยเฉพาะอนุมูลที่ไม่ชอบน้ำจะเข้าสู่โพรงของวงแหวนพี่เลี้ยง ในสภาพแวดล้อมเฉพาะของฟันผุเหล่านี้ การแจงนับของรูปแบบที่เป็นไปได้จะเกิดขึ้นจนกว่าจะพบเพียงรูปแบบเดียวที่เป็นประโยชน์มากที่สุดเท่านั้น (รูปที่ 1.27, B) การก่อตัวของโครงสร้างและการปลดปล่อยโปรตีนนั้นมาพร้อมกับการไฮโดรไลซิสของ ATP ในบริเวณเส้นศูนย์สูตร โดยทั่วไป การพับแบบขึ้นกับพี่เลี้ยงนั้นต้องการพลังงานจำนวนมาก
นอกเหนือจากการมีส่วนร่วมในการก่อตัวของโครงสร้างสามมิติของโปรตีนและการฟื้นฟูโปรตีนที่แปลงสภาพบางส่วนแล้ว พี่เลี้ยงยังจำเป็นสำหรับกระบวนการพื้นฐานเช่นการรวมตัวของโปรตีนโอลิโกเมอริก การรับรู้และการขนส่งโปรตีนที่แปลงสภาพเป็นไลโซโซม การขนส่งโปรตีน ข้ามเยื่อหุ้มและการมีส่วนร่วมในการควบคุมกิจกรรมของโปรตีนเชิงซ้อน
หัวข้อ 1.6. ความหลากหลายของโปรตีน กลุ่มโปรตีนในตัวอย่างของอิมมูโนโกลบูลิน
1. โปรตีนมีบทบาทชี้ขาดในชีวิตของเซลล์แต่ละเซลล์และสิ่งมีชีวิตหลายเซลล์ทั้งหมด และหน้าที่ของโปรตีนนั้นมีความหลากหลายอย่างน่าประหลาดใจ สิ่งนี้ถูกกำหนดโดยลักษณะเฉพาะของโครงสร้างหลักและโครงสร้างของโปรตีน โครงสร้างเฉพาะของศูนย์แอคทีฟ และความสามารถในการจับลิแกนด์จำเพาะ
มีเพียงส่วนเล็ก ๆ ของสายโซ่เปปไทด์ที่เป็นไปได้ทั้งหมดเท่านั้นที่สามารถนำมาใช้โครงสร้างเชิงพื้นที่ที่มั่นคงได้ ข้างมาก
ของพวกมันสามารถแปลงร่างได้หลายอย่างด้วยพลังงานกิ๊บส์ที่ใกล้เคียงกัน แต่ด้วย คุณสมบัติต่างๆ. โครงสร้างหลักของโปรตีนที่รู้จักมากที่สุด ซึ่งเลือกโดยวิวัฒนาการทางชีววิทยา ให้ความเสถียรที่ยอดเยี่ยมของโครงสร้างแบบใดรูปแบบหนึ่ง ซึ่งกำหนดคุณลักษณะของการทำงานของโปรตีนนี้
2. ตระกูลโปรตีนภายในสปีชีส์ทางชีววิทยาเดียวกัน การแทนที่ของเรซิดิวกรดอะมิโนสามารถนำไปสู่การเกิดขึ้นของโปรตีนที่แตกต่างกันซึ่งทำหน้าที่ที่เกี่ยวข้องกันและมีลำดับกรดอะมิโนที่คล้ายคลึงกัน โปรตีนที่เกี่ยวข้องดังกล่าวมีโครงสร้างที่คล้ายกันอย่างน่าทึ่ง: จำนวนและการจัดเรียงของ α-เฮลิซและ/หรือโครงสร้าง β และการหมุนและการพับส่วนใหญ่ของสายโพลีเปปไทด์มีความคล้ายคลึงหรือเหมือนกัน โปรตีนที่มีบริเวณคล้ายคลึงกันของสายพอลิเปปไทด์ โครงสร้างที่คล้ายกัน และหน้าที่ที่เกี่ยวข้องจะถูกแยกออกเป็นตระกูลโปรตีน ตัวอย่างของตระกูลโปรตีน: โปรตีนซีรีน, ตระกูลอิมมูโนโกลบูลิน, ตระกูลไมโอโกลบิน
โปรตีนซีรีน- ตระกูลโปรตีนที่ทำหน้าที่ของเอนไซม์โปรตีโอไลติก เหล่านี้รวมถึงเอนไซม์ย่อยอาหาร - ไคโมทริปซิน, ทริปซิน, อีลาสเทสและปัจจัยการแข็งตัวของเลือด โปรตีนเหล่านี้มีกรดอะมิโนเหมือนกัน 40% และมีโครงสร้างคล้ายกันมาก (รูปที่ 1.29)

ข้าว. 1.29. โครงสร้างเชิงพื้นที่ของอีลาสเทส (A) และไคโมทริปซิน (B)
การแทนที่กรดอะมิโนบางตัวทำให้เกิดการเปลี่ยนแปลงในความจำเพาะของซับสเตรตของโปรตีนเหล่านี้และการเกิดขึ้นของความหลากหลายเชิงฟังก์ชันภายในแฟมิลี
3. ครอบครัวของอิมมูโนโกลบูลินในการทำงานของระบบภูมิคุ้มกัน บทบาทที่ยิ่งใหญ่เล่นโปรตีนของอิมมูโนโกลบูลิน superfamily ซึ่งรวมถึงสามตระกูลของโปรตีน:
แอนติบอดี (อิมมูโนโกลบูลิน);
ตัวรับ T-lymphocyte;
โปรตีนของคอมเพล็กซ์ histocompatibility ที่สำคัญ - คลาส MHC 1 และ 2 (คอมเพล็กซ์ Histocompatibility หลัก)
โปรตีนทั้งหมดเหล่านี้มีโครงสร้างโดเมน ประกอบด้วยโดเมนคล้ายภูมิคุ้มกันที่คล้ายคลึงกันและทำหน้าที่คล้ายคลึงกัน: พวกมันมีปฏิสัมพันธ์กับโครงสร้างแปลกปลอมไม่ว่าจะละลายในเลือด น้ำเหลืองหรือของเหลวระหว่างเซลล์ (แอนติบอดี) หรืออยู่บนพื้นผิวของเซลล์ (ของตัวเองหรือ ต่างชาติ).
4. แอนติบอดี- โปรตีนจำเพาะที่ผลิตโดย B-lymphocytes เพื่อตอบสนองต่อการบริโภคโครงสร้างแปลกปลอมที่เรียกว่า แอนติเจน
คุณสมบัติของโครงสร้างของแอนติบอดี
โมเลกุลแอนติบอดีที่ง่ายที่สุดประกอบด้วยสายโพลีเปปไทด์สี่สาย: สายเบาที่เหมือนกันสองสาย - L มีกรดอะมิโนประมาณ 220 ตัวและสายหนักเหมือนกันสองสาย - H ซึ่งประกอบด้วยกรดอะมิโน 440-700 ตัว สายโซ่ทั้งสี่สายในโมเลกุลแอนติบอดีเชื่อมต่อกันด้วยพันธะที่ไม่ใช่โควาเลนต์จำนวนมากและพันธะไดซัลไฟด์สี่พันธะ (รูปที่ 1.30)
สายเบาของแอนติบอดีประกอบด้วยสองโดเมน: ตัวแปร (VL) ที่อยู่ในบริเวณปลาย N ของสายพอลิเปปไทด์ และค่าคงที่ (CL) ซึ่งตั้งอยู่ที่ปลาย C โซ่หนักโดยทั่วไปมีสี่โดเมน: หนึ่งตัวแปร (VH) ที่ปลาย N และสามค่าคงที่ (CH1, CH2, CH3) (ดูรูปที่ 1.30) โดเมนอิมมูโนโกลบูลินแต่ละโดเมนมีโครงสร้างเสริม β-pleated โดยที่ซิสเตอีนตกค้างสองตัวเชื่อมโยงกันด้วยพันธะไดซัลไฟด์
ระหว่างโดเมนคงที่ CH1 และ CH2 สองโดเมนมีบริเวณที่มีโพรลีนเรซิดิวจำนวนมาก ซึ่งป้องกันการก่อตัวของโครงสร้างทุติยภูมิและอันตรกิริยาของสาย H ที่อยู่ใกล้เคียงในส่วนนี้ บริเวณบานพับนี้ให้ความยืดหยุ่นของโมเลกุลแอนติบอดี ระหว่างโดเมนที่แปรผันได้ของสายหนักและสายเบาเป็นตำแหน่งซึ่งจับแอนติเจนที่เหมือนกันสองแห่ง (ตำแหน่งที่ออกฤทธิ์สำหรับแอนติเจนในการจับ) ดังนั้นแอนติบอดีดังกล่าวจึงมักถูกเรียกว่า ไบวาเลนท์การจับของแอนติเจนกับแอนติบอดีไม่เกี่ยวข้องกับลำดับกรดอะมิโนทั้งหมดของบริเวณที่แปรผันได้ของทั้งสองสาย แต่มีกรดอะมิโนเพียง 20-30 ตัวเท่านั้นที่อยู่ในบริเวณที่แปรผันได้สูงของแต่ละสาย เป็นพื้นที่เหล่านี้ที่กำหนดความสามารถเฉพาะของแอนติบอดีแต่ละประเภทในการโต้ตอบกับแอนติเจนเสริมที่สอดคล้องกัน
แอนติบอดีเป็นหนึ่งในแนวป้องกันของร่างกายจากการบุกรุกสิ่งมีชีวิตต่างประเทศ การทำงานของพวกมันสามารถแบ่งออกเป็นสองขั้นตอน: ระยะแรกคือการรับรู้และการผูกมัดของแอนติเจนบนพื้นผิวของสิ่งมีชีวิตแปลกปลอม ซึ่งเป็นไปได้เนื่องจากการมีอยู่ของตำแหน่งที่จับกับแอนติเจนในโครงสร้างแอนติบอดี ขั้นตอนที่สองคือการเริ่มต้นกระบวนการปิดการใช้งานและการทำลายแอนติเจน ความจำเพาะของระยะที่สองขึ้นอยู่กับประเภทของแอนติบอดี โซ่หนักมีห้าคลาสที่แตกต่างกันในโครงสร้างของโดเมนคงที่: α, δ, ε, γ และ μ ตามประเภทของอิมมูโนโกลบูลินห้าคลาส: A, D, E, G และ M
ลักษณะโครงสร้างของสายหนักทำให้บริเวณบานพับและบริเวณปลาย C ของสายหนักมีลักษณะโครงสร้างของแต่ละชั้น เมื่อแอนติเจนจับกับแอนติบอดี การเปลี่ยนแปลงเชิงโครงสร้างในโดเมนคงที่กำหนดวิถีสำหรับการกำจัดแอนติเจน
 ข้าว. 1. 30. โครงสร้างโดเมนของ IgG
ข้าว. 1. 30. โครงสร้างโดเมนของ IgG
อิมมูโนโกลบูลิน M
อิมมูโนโกลบูลิน M มีสองรูปแบบ
แบบฟอร์มโมโนเมอร์- แอนติบอดีประเภทที่ 1 ที่ผลิตโดย B-lymphocyte ที่กำลังพัฒนา ต่อจากนั้น บีเซลล์จำนวนมากเปลี่ยนไปผลิตแอนติบอดีประเภทอื่น แต่มีตำแหน่งจับแอนติเจนเดียวกัน IgM ถูกรวมเข้าไปในเมมเบรนและทำหน้าที่เป็นตัวรับที่รับรู้แอนติเจน การรวมตัวของ IgM เข้าไปในเยื่อหุ้มเซลล์เป็นไปได้เนื่องจากการมีอยู่ของกรดอะมิโนที่ไม่ชอบน้ำ 25 ตัวในส่วนหางของบริเวณ
รูปแบบสารคัดหลั่งของ IgMประกอบด้วยหน่วยย่อยโมโนเมอร์ห้ายูนิตที่เชื่อมโยงซึ่งกันและกันโดยพันธะไดซัลไฟด์และสาย J โพลีเปปไทด์เพิ่มเติม (รูปที่ 1.31) โมโนเมอร์สายหนักในแบบฟอร์มนี้ไม่มีหางที่ไม่ชอบน้ำ เพนทาเมอร์มีจุดจับแอนติเจน 10 ตำแหน่ง ดังนั้นจึงมีประสิทธิภาพในการจดจำและกำจัดแอนติเจนที่เข้าสู่ร่างกายเป็นครั้งแรก รูปแบบสารคัดหลั่งของ IgM เป็นแอนติบอดีประเภทหลักที่หลั่งเข้าสู่กระแสเลือดระหว่างการตอบสนองทางภูมิคุ้มกันเบื้องต้น การจับของ IgM กับแอนติเจนจะเปลี่ยนโครงสร้างของ IgM และชักนำให้เกิดการจับกับส่วนประกอบโปรตีนแรกของระบบคอมพลีเมนต์ (ระบบคอมพลีเมนต์คือชุดของโปรตีนที่เกี่ยวข้องกับการทำลายแอนติเจน) และการกระตุ้นระบบนี้ หากแอนติเจนอยู่บนพื้นผิวของจุลินทรีย์ระบบเสริมจะทำให้เกิดการละเมิดความสมบูรณ์ เยื่อหุ้มเซลล์และการตายของเซลล์แบคทีเรีย
อิมมูโนโกลบูลิน G
ในแง่เชิงปริมาณ อิมมูโนโกลบูลินประเภทนี้มีอิทธิพลเหนือในเลือด (75% ของ Ig ทั้งหมด) IgG - โมโนเมอร์ซึ่งเป็นแอนติบอดีระดับหลักที่หลั่งเข้าสู่กระแสเลือดระหว่างการตอบสนองของภูมิคุ้มกันทุติยภูมิ หลังจากการทำงานร่วมกันของ IgG กับแอนติเจนบนพื้นผิวของจุลินทรีย์ คอมเพล็กซ์แอนติเจน-แอนติบอดีสามารถจับและกระตุ้นโปรตีนของระบบเสริม หรือสามารถโต้ตอบกับตัวรับจำเพาะบนมาโครฟาจและนิวโทรฟิล ปฏิสัมพันธ์กับฟาโกไซต์
 ข้าว. 1.31. โครงสร้างของสารคัดหลั่งของ IgM
ข้าว. 1.31. โครงสร้างของสารคัดหลั่งของ IgM
ต่อการดูดซึมของสารเชิงซ้อนของแอนติเจนและแอนติบอดีและการทำลายของพวกมันในฟาโกโซมของเซลล์ IgG เป็นแอนติบอดีประเภทเดียวที่สามารถข้ามอุปสรรครกและป้องกันทารกในครรภ์จากการติดเชื้อในครรภ์ได้
อิมมูโนโกลบูลิน A
แอนติบอดีประเภทหลักมีอยู่ในสารคัดหลั่ง (นม น้ำลาย สารคัดหลั่งจากทางเดินหายใจและลำไส้) IgA ถูกคัดหลั่งออกมาในรูปแบบไดเมอร์เป็นหลัก โดยที่โมโนเมอร์เชื่อมโยงถึงกันผ่านสายโซ่ J เพิ่มเติม (รูปที่ 1.32)
IgA ไม่มีปฏิกิริยากับระบบเสริมและเซลล์ฟาโกไซติก แต่โดยการจับกับจุลินทรีย์ แอนติบอดีจะป้องกันไม่ให้พวกมันเกาะติดกับเซลล์เยื่อบุผิวและแทรกซึมเข้าสู่ร่างกาย
อิมมูโนโกลบูลิน E
อิมมูโนโกลบูลิน E แสดงโดยโมโนเมอร์ซึ่งมี ε-chains หนัก เช่นเดียวกับ μ-chains ของอิมมูโนโกลบูลิน M หนึ่งตัวแปรและโดเมนคงที่สี่โดเมน IgE หลังการหลั่งจะผูกมัดตัวเอง
 ข้าว. 1.32. โครงสร้างของ IgA
ข้าว. 1.32. โครงสร้างของ IgA
บริเวณปลาย C ที่มีตัวรับที่สอดคล้องกันบนพื้นผิวของแมสต์เซลล์และเบสโซฟิล เป็นผลให้พวกมันกลายเป็นตัวรับแอนติเจนบนพื้นผิวของเซลล์เหล่านี้ (รูปที่ 1.33)
 ข้าว. 1.33. ปฏิกิริยาของ IgE กับแอนติเจนบนพื้นผิวของเซลล์เสา
ข้าว. 1.33. ปฏิกิริยาของ IgE กับแอนติเจนบนพื้นผิวของเซลล์เสา
หลังจากที่แอนติเจนติดอยู่กับตำแหน่ง IgE ที่จับกับแอนติเจนแล้ว เซลล์จะได้รับสัญญาณเพื่อหลั่งสารออกฤทธิ์ทางชีวภาพ (ฮีสตามีน, เซโรโทนิน) ซึ่งส่วนใหญ่รับผิดชอบในการพัฒนาปฏิกิริยาการอักเสบและสำหรับการแสดงปฏิกิริยาการแพ้เช่น โรคหอบหืดลมพิษไข้ละอองฟาง
อิมมูโนโกลบูลิน D
อิมมูโนโกลบูลินดีพบได้ในซีรั่มมาก ในปริมาณที่น้อย, พวกมันเป็นโมโนเมอร์ โซ่ δ หนักมีหนึ่งตัวแปรและโดเมนคงที่สามโดเมน IgD ทำหน้าที่เป็นตัวรับ B-lymphocytes ส่วนหน้าที่อื่นๆ ยังไม่ทราบ อันตรกิริยาของแอนติเจนจำเพาะกับตัวรับบนพื้นผิวของ B-lymphocytes (IgD) นำไปสู่การส่งสัญญาณเหล่านี้ไปยังเซลล์และการกระตุ้นกลไกที่รับประกันการสืบพันธุ์ของโคลนของเซลล์เม็ดเลือดขาวนี้
หัวข้อ 1.7. คุณสมบัติทางกายภาพและเคมีของโปรตีนและวิธีการแยกออก
1. โปรตีนแต่ละตัวมีคุณสมบัติทางเคมีกายภาพแตกต่างกัน:
รูปร่างของโมเลกุล
น้ำหนักโมเลกุล
ประจุทั้งหมด ค่าที่ขึ้นอยู่กับอัตราส่วนของกลุ่มประจุลบและประจุบวกของกรดอะมิโน
อัตราส่วนของอนุมูลของกรดอะมิโนแบบมีขั้วและแบบไม่มีขั้วบนผิวของโมเลกุล
องศาของความต้านทานต่อสารเปลี่ยนสภาพต่างๆ
2. ความสามารถในการละลายของโปรตีนขึ้นอยู่กับเกี่ยวกับคุณสมบัติของโปรตีนที่ระบุไว้ข้างต้น เช่นเดียวกับองค์ประกอบของตัวกลางที่โปรตีนละลาย (ค่า pH องค์ประกอบของเกลือ อุณหภูมิ การปรากฏตัวของสารอินทรีย์อื่นๆ ที่สามารถโต้ตอบกับโปรตีนได้) ขนาดของประจุของโมเลกุลโปรตีนเป็นหนึ่งในปัจจัยที่ส่งผลต่อความสามารถในการละลายของพวกมัน เมื่อประจุหายไปที่จุดไอโซอิเล็กทริก โปรตีนจะรวมตัวกันและตกตะกอนได้ง่ายขึ้น โดยเฉพาะอย่างยิ่งสำหรับโปรตีนที่เสียสภาพซึ่งมีอนุมูลของกรดอะมิโนที่ไม่ชอบน้ำบนผิว
บนพื้นผิวของโมเลกุลโปรตีนนั้นมีทั้งอนุมูลของกรดอะมิโนที่มีประจุบวกและลบ จำนวนของกลุ่มเหล่านี้และด้วยเหตุนี้ประจุทั้งหมดของโปรตีนจึงขึ้นอยู่กับ pH ของตัวกลางนั่นคือ อัตราส่วนความเข้มข้นของกลุ่ม H + - และ OH ในสภาพแวดล้อมที่เป็นกรดการเพิ่มความเข้มข้นของ H+ นำไปสู่การปราบปรามการแยกตัวของกลุ่มคาร์บอกซิล -COO - + H+ > -COOH และการลดลงของประจุลบของโปรตีน ในสภาพแวดล้อมที่เป็นด่างการจับกันของ OH - โปรตอนส่วนเกินที่เกิดขึ้นในระหว่างการแยกตัวของหมู่อะมิโน -NH 3 + + OH - - NH 2 + H 2 O กับการก่อตัวของน้ำทำให้ประจุบวกของโปรตีนลดลง ค่า pH ที่โปรตีนมีประจุสุทธิเป็นศูนย์เรียกว่า จุดไอโซอิเล็กทริก (IEP)ใน IET จำนวนกลุ่มที่มีประจุบวกและลบจะเท่ากัน กล่าวคือ โปรตีนอยู่ในสถานะไอโซอิเล็กทริก
3. การแยกโปรตีนแต่ละตัวคุณสมบัติของโครงสร้างและการทำงานของร่างกายขึ้นอยู่กับชุดของโปรตีนที่สังเคราะห์ขึ้น การศึกษาโครงสร้างและคุณสมบัติของโปรตีนเป็นไปไม่ได้หากปราศจากการแยกตัวออกจากเซลล์และทำให้บริสุทธิ์จากโปรตีนอื่นๆ และโมเลกุลอินทรีย์ ขั้นตอนของการแยกและทำให้บริสุทธิ์ของโปรตีนแต่ละตัว:
การทำลายเซลล์ของเนื้อเยื่อที่ศึกษาและได้รับโฮโมจีเนต
การแยกโฮโมจีเนตออกเป็นเศษส่วนการหมุนเหวี่ยงเพื่อให้ได้นิวเคลียส ไมโทคอนเดรีย ไซโตซอลหรือส่วนอื่นๆ ที่มีโปรตีนที่ต้องการ
การเปลี่ยนสภาพความร้อนแบบเลือกได้- การให้ความร้อนในระยะสั้นของสารละลายโปรตีน ซึ่งสามารถขจัดสิ่งเจือปนโปรตีนบางส่วนออกได้ (ในกรณีที่โปรตีนค่อนข้างคงตัวทางความร้อน)
เกลือออกโปรตีนต่าง ๆ ตกตะกอนที่ความเข้มข้นของเกลือต่างกันในสารละลาย โดยการค่อยๆ เพิ่มความเข้มข้นของเกลือ เป็นไปได้ที่จะได้เศษส่วนจำนวนหนึ่งซึ่งมีเนื้อหาเด่นของโปรตีนที่หลั่งออกมาในหนึ่งในนั้น การแยกส่วนโปรตีนที่ใช้บ่อยที่สุดคือแอมโมเนียมซัลเฟต โปรตีนที่มีความสามารถในการละลายต่ำที่สุดตกตะกอนที่ความเข้มข้นของเกลือต่ำ
กรองเจล- วิธีการกรองโมเลกุลผ่านเม็ด Sephadex ที่บวม (สายโซ่เดกซ์ตรอนสามมิติที่มีรูพรุน) อัตราการผ่านของโปรตีนผ่านคอลัมน์ที่เต็มไปด้วย Sephadex จะขึ้นอยู่กับน้ำหนักโมเลกุลของพวกมัน: ยิ่งโมเลกุลของโปรตีนมีมวลน้อยเท่าไร พวกมันก็ยิ่งเจาะเข้าไปในแกรนูลได้ง่ายขึ้นและอยู่ที่นั่นนานขึ้น ยิ่งมวลมากขึ้นเท่านั้น พวกมันก็จะหลุดออกจาก คอลัมน์.
Ultracentrifugation- วิธีการประกอบด้วยการใส่โปรตีนในหลอดหมุนเหวี่ยงในโรเตอร์ของเครื่องหมุนเหวี่ยงอัลตร้าโซนิค เมื่อโรเตอร์หมุน อัตราการตกตะกอนของโปรตีนจะแปรผันตามน้ำหนักโมเลกุล: เศษส่วนของโปรตีนที่หนักกว่าจะอยู่ใกล้ก้นหลอดมากขึ้น และส่วนที่เบากว่าจะอยู่ใกล้พื้นผิวมากขึ้น
อิเล็กโตรโฟรีซิส- วิธีการขึ้นอยู่กับความแตกต่างของความเร็วของการเคลื่อนที่ของโปรตีนในสนามไฟฟ้า ค่านี้เป็นสัดส่วนกับประจุของโปรตีน โปรตีนอิเล็กโตรโฟรีซิสดำเนินการบนกระดาษ (ในกรณีนี้ ความเร็วของโปรตีนเป็นสัดส่วนกับประจุเท่านั้น) หรือในเจลโพลีอะคริลาไมด์ที่มีขนาดรูพรุนที่แน่นอน (ความเร็วของโปรตีนเป็นสัดส่วนกับประจุและน้ำหนักโมเลกุล)
โครมาโตกราฟีแลกเปลี่ยนไอออน- วิธีการแยกส่วนตามการจับของกลุ่มโปรตีนที่แตกตัวเป็นไอออนกับกลุ่มของเรซินแลกเปลี่ยนไอออนที่มีประจุตรงข้าม (วัสดุพอลิเมอร์ที่ไม่ละลายน้ำ) ความแข็งแรงในการยึดเกาะของโปรตีนกับเรซินนั้นแปรผันตามประจุของโปรตีน โปรตีนที่ดูดซับบนพอลิเมอร์แลกเปลี่ยนไอออนสามารถชะล้างออกได้ด้วยความเข้มข้นของสารละลาย NaCl ที่เพิ่มขึ้น ยิ่งประจุโปรตีนต่ำเท่าใด ความเข้มข้นของ NaCl ก็ยิ่งต่ำลงเพื่อล้างโปรตีนที่เกี่ยวข้องกับกลุ่มไอออนิกของเรซินออกไป
โครมาโตกราฟีสัมพันธ์- วิธีเฉพาะเจาะจงที่สุดในการแยกโปรตีนแต่ละชนิด ลิแกนด์ของโปรตีนติดอยู่กับพอลิเมอร์เฉื่อยแบบโควาเลนต์ เมื่อสารละลายโปรตีนถูกส่งผ่านคอลัมน์ที่มีพอลิเมอร์ เนื่องจากการยึดเกาะของโปรตีนกับลิแกนด์ เฉพาะโปรตีนที่จำเพาะสำหรับลิแกนด์นี้เท่านั้นที่ถูกดูดซับบนคอลัมน์
ฟอกไต- วิธีการที่ใช้ในการกำจัดสารประกอบที่มีน้ำหนักโมเลกุลต่ำออกจากสารละลายของโปรตีนที่แยกได้ วิธีการนี้ขึ้นอยู่กับความสามารถของโปรตีนที่จะผ่านเมมเบรนแบบกึ่งซึมผ่านได้ ตรงกันข้ามกับสารที่มีน้ำหนักโมเลกุลต่ำ ใช้เพื่อทำให้โปรตีนบริสุทธิ์จากสิ่งเจือปนที่มีโมเลกุลต่ำ เช่น จากเกลือหลังจากเกลือออก
การมอบหมายงานนอกหลักสูตร
1. กรอกตาราง. 1.4.
ตารางที่ 1.4. การวิเคราะห์เปรียบเทียบโครงสร้างและหน้าที่ของโปรตีนที่เกี่ยวข้อง - myoglobin และ hemoglobin
ก) จำโครงสร้างของศูนย์ที่ใช้งานอยู่ Mb และ Hb อนุมูลที่ไม่ชอบน้ำของกรดอะมิโนมีบทบาทอย่างไรในการก่อตัวของศูนย์กลางของโปรตีนเหล่านี้? อธิบายโครงสร้างของศูนย์แอคทีฟ Mb และ Hb และกลไกการยึดเกาะของลิแกนด์ สารตกค้าง F 8 และ E 7 ของเขามีบทบาทอย่างไรในการทำงานของไซต์แอคทีฟ Mv และ Hv
ข) คุณสมบัติใหม่อะไรเมื่อเทียบกับโมโนเมอร์ myoglobin ที่มีโปรตีนโอลิโกเมอร์ที่เกี่ยวข้องอย่างใกล้ชิดคือเฮโมโกลบิน? อธิบายบทบาทของการเปลี่ยนแปลงร่วมกันในโครงสร้างของโปรโตเมอร์ในโมเลกุลของเฮโมโกลบิน ผลของ CO 2 และความเข้มข้นของโปรตอนต่อสัมพรรคภาพระหว่างฮีโมโกลบินกับออกซิเจน และบทบาทของ 2,3-BPG ในการควบคุมอัลโลสเตอริกของฟังก์ชัน Hb
2. อธิบายลักษณะของโมเลกุลพี่เลี้ยงโดยให้ความสนใจกับความสัมพันธ์ระหว่างโครงสร้างและหน้าที่ของพวกมัน
3. โปรตีนอะไรถูกจัดกลุ่มเป็นครอบครัว? โดยใช้ตัวอย่างของตระกูลอิมมูโนโกลบูลิน กำหนดลักษณะโครงสร้างที่คล้ายคลึงกันและหน้าที่ที่เกี่ยวข้องของโปรตีนในตระกูลนี้
4. บ่อยครั้ง โปรตีนบริสุทธิ์แต่ละชนิดจำเป็นสำหรับวัตถุประสงค์ทางชีวเคมีและทางการแพทย์ อธิบายคุณสมบัติทางเคมีกายภาพของโปรตีนที่ใช้สำหรับการแยกและการทำให้บริสุทธิ์
งานเพื่อการควบคุมตนเอง
1. เลือกคำตอบที่ถูกต้อง.
หน้าที่ของเฮโมโกลบิน:
ก. O 2 ลำเลียงจากปอดไปยังเนื้อเยื่อ B. H + ขนส่งจากเนื้อเยื่อไปยังปอด
ข. รักษาค่า pH ของเลือดให้คงที่ ง. ขนส่ง CO2 จากปอดไปยังเนื้อเยื่อ
ง. การขนส่ง CO 2 จากเนื้อเยื่อไปยังปอด
2. เลือกคำตอบที่ถูกต้อง. ลิแกนด์α -Hb protomer คือ:ก. ฮิเมะ
ข. ออกซิเจน
B. CO D. 2,3-BPG
D. β-โปรโตเมอร์
3. เลือกคำตอบที่ถูกต้อง.
เฮโมโกลบินแตกต่างจาก myoglobin:
ก. มีโครงสร้างเป็นสี่ส่วน
B. โครงสร้างทุติยภูมิแสดงโดย α-helices . เท่านั้น
B. หมายถึงโปรตีนที่ซับซ้อน
ง. โต้ตอบกับลิแกนด์อัลโลสเตอริก ง. ผูกพันกับฮีมอย่างโควาเลนต์
4. เลือกคำตอบที่ถูกต้อง.
ความสัมพันธ์ของ Hb สำหรับ O 2 ลดลง:
ก. เมื่อ O 2 โมเลกุลเกาะติดกัน B. เมื่อโมเลกุล O 2 ตัวหนึ่งถูกกำจัดออก
B. เมื่อโต้ตอบกับ 2,3-BPG
ง. เมื่อติดโปรโตเมอร์ H + D เมื่อความเข้มข้นของ 2,3-BPG ลดลง
5. ตั้งค่าการแข่งขัน
สำหรับประเภท Hb เป็นลักษณะเฉพาะ:
A. สร้างมวลรวมของไฟบริลลาร์ในรูปแบบดีออกซี B. ประกอบด้วยสายโซ่ α- สองตัวและสายโซ่ δ สองสาย
B. รูปแบบเด่นของ Hb ในเม็ดเลือดแดงของผู้ใหญ่ D. ประกอบด้วย heme ที่มี Fe + 3 ในศูนย์ที่ใช้งานอยู่
D. ประกอบด้วยสาย α- สองสายและสาย γ สองสาย 1. HvA 2
6. ตั้งค่าการแข่งขัน
แกนด์ Hb:
A. ผูกกับ Hb ที่ allosteric center
B. มีความสัมพันธ์ที่สูงมากสำหรับไซต์ที่ใช้งานอยู่ Hb
B. เข้าร่วมเพิ่มความสัมพันธ์ของ Hb กับ O 2 D. ออกซิไดซ์ Fe + 2 ถึง Fe + 3
ง. สร้างพันธะโควาเลนต์กับ hysF8
7. เลือกคำตอบที่ถูกต้อง.
พี่เลี้ยง:
ก. โปรตีนมีอยู่ในทุกส่วนของเซลล์
B. การสังเคราะห์ได้รับการปรับปรุงภายใต้อิทธิพลที่กดดัน
B. มีส่วนร่วมในการไฮโดรไลซิสของโปรตีนที่ทำให้เสียสภาพ
ง. มีส่วนร่วมในการรักษารูปแบบดั้งเดิมของโปรตีน
ง. สร้างออร์แกเนลล์ที่มีโปรตีนก่อตัวขึ้น
8. แมทช์ อิมมูโนโกลบูลิน:
ก. รูปแบบสารคัดหลั่งคือเพนทาเมริก
B. ชั้น Ig ที่ข้ามชั้นรก
B. Ig - ตัวรับเซลล์แมสต์
D. คลาสหลักของ Ig มีอยู่ในการหลั่งของเซลล์เยื่อบุผิว ดี. บี-ลิมโฟไซต์ รีเซพเตอร์ ซึ่งกระตุ้นการสืบพันธุ์ของเซลล์
9. เลือกคำตอบที่ถูกต้อง.
อิมมูโนโกลบูลินอี:
ก. ผลิตโดยแมคโครฟาจ ข. มีโซ่ ε หนัก
B. ฝังอยู่ในเยื่อหุ้มเซลล์ T-lymphocytes
ง. ทำหน้าที่เป็นตัวรับเมมเบรนสำหรับแอนติเจนบนแมสต์เซลล์และเบสโซฟิล
D. รับผิดชอบในการสำแดงอาการแพ้
10. เลือกคำตอบที่ถูกต้อง.
วิธีการแยกโปรตีนขึ้นอยู่กับความแตกต่างของน้ำหนักโมเลกุล:
ก. การกรองแบบเจล
B. การหมุนเหวี่ยงด้วยคลื่นอัลตร้าโซนิค
B. โพลีอะคริลาไมด์เจลอิเล็กโตรโฟรีซิส D. โครมาโตกราฟีแลกเปลี่ยนไอออน
ง. Affinity chromatography
11. เลือกคำตอบที่ถูกต้อง.
วิธีการแยกโปรตีนขึ้นอยู่กับความแตกต่างในการละลายในน้ำ:
ก. การกรองเจล ข. การทำเกลือออก
B. โครมาโตกราฟีแลกเปลี่ยนไอออน ง. โครมาโตกราฟีสัมพันธ์
E. โพลีอะคริลาไมด์เจลอิเล็กโทรโฟรีซิส
มาตรฐานของคำตอบ "ภารกิจเพื่อการควบคุมตนเอง"
1. A, B, C, D
2. A, B, C, D
5. 1-B, 2-A, 3-G
6. 1-C, 2-B, 3-A
7. A, B, D, D
8. 1-G; 2-B, 3-C
ข้อกำหนดและแนวคิดพื้นฐาน
1. โปรตีนโอลิโกเมอร์ โปรโตเมอร์ โครงสร้างควอเทอร์นารีของโปรตีน
2. การเปลี่ยนแปลงของสหกรณ์ในรูปแบบโปรโตเมอร์
3. เอฟเฟกต์บอร์
4. การควบคุม allosteric ของฟังก์ชันโปรตีน allosteric center และ allosteric effector
5. โมเลกุลพี่เลี้ยง โปรตีนช็อกความร้อน
6. กลุ่มโปรตีน (ซีรีนโปรตีเอส, อิมมูโนโกลบูลิน)
7. IgM-, G-, E-, A- การเชื่อมต่อของโครงสร้างพร้อมฟังก์ชัน
8. ประจุรวมของโปรตีน จุดไอโซอิเล็กทริกของโปรตีน
9. อิเล็กโทรโฟรีซิส
10. เกลือออก
11. การกรองเจล
12. โครมาโตกราฟีแลกเปลี่ยนไอออน
13. Ultracentrifugation
14. โครมาโตกราฟีสัมพันธ์
15. อิเล็กโตรโฟรีซิสโปรตีนในพลาสมา
งานสำหรับงานออดิชั่น
1. เปรียบเทียบการพึ่งพาระดับความอิ่มตัวของฮีโมโกลบิน (Hb) และไมโอโกลบิน (Mb) กับออกซิเจนกับความดันบางส่วนในเนื้อเยื่อ
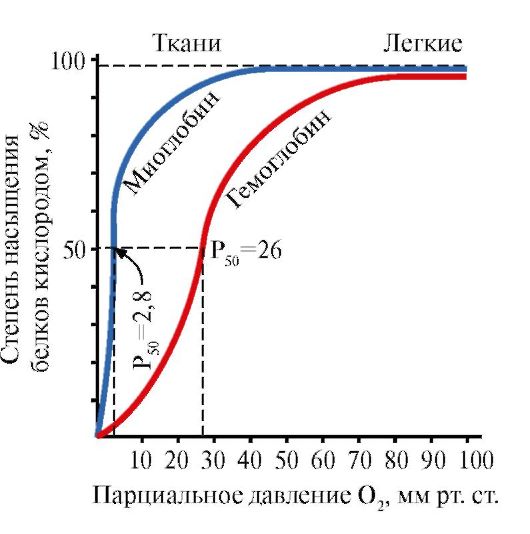 ข้าว. 1.34. การพึ่งพาความอิ่มตัวของ MV และHBออกซิเจนจากความดันบางส่วน
ข้าว. 1.34. การพึ่งพาความอิ่มตัวของ MV และHBออกซิเจนจากความดันบางส่วน
โปรดทราบว่ารูปร่างของเส้นโค้งความอิ่มตัวของออกซิเจนในโปรตีนนั้นแตกต่างกัน: สำหรับ myoglobin - อติพจน์ สำหรับเฮโมโกลบิน - รูปร่างซิกมอยด์
1. เปรียบเทียบค่าความดันบางส่วนของออกซิเจนที่ Mb และ Hb อิ่มตัวด้วย O 2 โดย 50% โปรตีนใดต่อไปนี้มีความสัมพันธ์ใกล้ชิดกับ O 2 มากกว่า
2. ลักษณะโครงสร้างของ MB ใดที่กำหนดความสัมพันธ์สูงสำหรับ O 2 ?
3. ลักษณะโครงสร้างใดของ Hb ที่อนุญาตให้ปล่อย O 2 ในเส้นเลือดฝอยของเนื้อเยื่อที่พัก (ที่ความดันบางส่วนที่ค่อนข้างสูงของ O 2) และเพิ่มผลตอบแทนของกล้ามเนื้อทำงานอย่างรวดเร็ว คุณสมบัติใดของโปรตีนโอลิโกเมอร์ที่ให้ผลนี้?
4. คำนวณปริมาณ O 2 (เป็น%) ที่ให้ออกซิเจนเฮโมโกลบินแก่กล้ามเนื้อพักและทำงานอย่างไร
5. หาข้อสรุปเกี่ยวกับความสัมพันธ์ระหว่างโครงสร้างโปรตีนและหน้าที่ของมัน
2. ปริมาณออกซิเจนที่ปล่อยออกมาจากเฮโมโกลบินในเส้นเลือดฝอยขึ้นอยู่กับความเข้มข้นของกระบวนการแคแทบอลิซึมในเนื้อเยื่อ (ผลของบอร์) การเปลี่ยนแปลงในการเผาผลาญเนื้อเยื่อควบคุมความสัมพันธ์ของ Hb สำหรับ O 2 ได้อย่างไร? ผลของ CO 2 และ H+ ต่อสัมพรรคภาพของ Hb กับ O 2
 1. อธิบายเอฟเฟกต์ของบอร์
1. อธิบายเอฟเฟกต์ของบอร์
2. กระบวนการที่แสดงในแผนภาพโฟลว์ไปในทิศทางใด:
ก) ในเส้นเลือดฝอยของปอด;
b) ในเส้นเลือดฝอยเนื้อเยื่อ?
3. ความสำคัญทางสรีรวิทยาของผลกระทบจากบอร์คืออะไร?
4. เหตุใดปฏิสัมพันธ์ของ Hb กับ H+ ที่ไซต์ที่ห่างไกลจาก heme จึงเปลี่ยนความสัมพันธ์ของโปรตีนสำหรับ O 2
3. ความสัมพันธ์ของ Hb กับ O 2 ขึ้นอยู่กับความเข้มข้นของลิแกนด์ 2,3-biphosphoglycerate ซึ่งเป็นตัวควบคุม allosteric ของความสัมพันธ์ของ Hb กับ O 2 เหตุใดปฏิสัมพันธ์ของลิแกนด์ที่ไซต์ระยะไกลจากไซต์ที่ทำงานอยู่จึงส่งผลต่อการทำงานของโปรตีน 2,3-BPG ควบคุมความสัมพันธ์ของ Hb สำหรับ O 2 อย่างไร ในการแก้ปัญหา ให้ตอบคำถามต่อไปนี้:
1. สังเคราะห์ 2,3-biphosphoglycerate (2,3-BPG) ที่ไหนและจากอะไร? เขียนสูตรระบุประจุของโมเลกุลนี้
2. BPG โต้ตอบกับเฮโมโกลบินรูปแบบใด (ออกซีหรือดีออกซี) และเพราะเหตุใด ปฏิสัมพันธ์เกิดขึ้นที่บริเวณใดของโมเลกุล Hb
3. กระบวนการที่แสดงในแผนภาพดำเนินการไปในทิศทางใด
ก) ในเส้นเลือดฝอยเนื้อเยื่อ;
b) ในเส้นเลือดฝอยของปอด?
4. ที่ไหนควรจะมีความเข้มข้นสูงสุดของคอมเพล็กซ์
Nv-2,3-BFG:
ก) ในเส้นเลือดฝอยของกล้ามเนื้อที่เหลือ
b) ในเส้นเลือดฝอยของกล้ามเนื้อทำงาน (สมมติว่า BPG มีความเข้มข้นเท่ากันในเม็ดเลือดแดง)?
5. ความสัมพันธ์ของ Hb ต่อออกซิเจนจะเปลี่ยนไปอย่างไรเมื่อบุคคลปรับตัวให้เข้ากับสภาวะระดับความสูง ถ้าความเข้มข้นของ BPG ในเม็ดเลือดแดงเพิ่มขึ้น? ความสำคัญทางสรีรวิทยาของปรากฏการณ์นี้คืออะไร?
4. การทำลาย 2,3-BPG ระหว่างการเก็บรักษาเลือดที่เก็บรักษาไว้จะขัดขวางการทำงานของ Hb ความสัมพันธ์ของ Hb กับ O 2 ในเลือดที่เก็บรักษาไว้จะเปลี่ยนไปอย่างไรหากความเข้มข้น 2,3-BPG ในเม็ดเลือดแดงลดลงจาก 8 เป็น 0.5 mmol/l เป็นไปได้หรือไม่ที่จะถ่ายเลือดดังกล่าวให้กับผู้ป่วยที่ป่วยหนักถ้าความเข้มข้น 2,3-BPG กลับคืนมาไม่เร็วกว่าสามวัน? เป็นไปได้ไหมที่จะฟื้นฟูการทำงานของเม็ดเลือดแดงโดยการเพิ่ม 2,3-BPG ในเลือด?
5. จำโครงสร้างของโมเลกุลอิมมูโนโกลบูลินที่ง่ายที่สุด อิมมูโนโกลบูลินมีบทบาทอย่างไรในระบบภูมิคุ้มกัน? ทำไม Igs มักถูกเรียกว่า bivalents? โครงสร้างของ Igs สัมพันธ์กับหน้าที่อย่างไร? (อธิบายโดยใช้ตัวอย่างคลาสของอิมมูโนโกลบูลิน)
คุณสมบัติทางกายภาพและเคมีของโปรตีนและวิธีการแยกโปรตีน
6. ประจุสุทธิของโปรตีนส่งผลต่อความสามารถในการละลายของมันอย่างไร?
ก) กำหนดประจุทั้งหมดของเปปไทด์ที่ pH 7
Ala-Glu-Tre-Pro-Asp-Liz-Cis
b) ประจุของเปปไทด์นี้จะเปลี่ยนแปลงอย่างไรที่ pH >7, pH<7, рН <<7?
c) จุดไอโซอิเล็กทริกของโปรตีน (IEP) คืออะไรและอยู่ในสภาพแวดล้อมใด
IET ของเปปไทด์นี้?
d) ค่า pH ใดที่จะสามารถละลายได้น้อยที่สุดของเปปไทด์นี้
7. ทำไมนมเปรี้ยวจึง "จับตัวเป็นก้อน" เมื่อต้ม (เช่น โปรตีนจากนมเคซีนตกตะกอน) ไม่เหมือนกับนมสด โมเลกุลของเคซีนในนมสดมีประจุเป็นลบ
8. การกรองเจลใช้เพื่อแยกโปรตีนแต่ละตัว ของผสมที่มีโปรตีน A, B, C ที่มีน้ำหนักโมเลกุลเท่ากับ 160,000, 80,000 และ 60,000 ตามลำดับ ถูกวิเคราะห์โดยการกรองแบบเจล (รูปที่ 1.35) เม็ดเจลบวมสามารถซึมเข้าไปในโปรตีนที่มีน้ำหนักโมเลกุลน้อยกว่า 70,000 ได้ หลักการใดที่สนับสนุนวิธีการแยกนี้ กราฟใดแสดงผลการแยกส่วนได้ถูกต้อง ระบุลำดับการปลดปล่อยโปรตีน A, B และ C จากคอลัมน์
 ข้าว. 1.35. ใช้วิธีกรองเจลเพื่อแยกโปรตีน
ข้าว. 1.35. ใช้วิธีกรองเจลเพื่อแยกโปรตีน
9. ในรูป 1.36, A แสดงไดอะแกรมของอิเล็กโตรโฟรีซิสบนกระดาษของโปรตีนในซีรัมในเลือดของบุคคลที่มีสุขภาพดี ปริมาณสัมพัทธ์ของเศษส่วนโปรตีนที่ได้จากวิธีนี้ ได้แก่ อัลบูมิน 54-58%, α 1 -โกลบูลิน 6-7%, α 2 -โกลบูลิน 8-9%, β-โกลบูลิน 13%, γ-โกลบูลิน 11-12% .
 ข้าว. 1.36 อิเล็กโตรโฟรีซิสบนกระดาษของโปรตีนในพลาสมาในเลือดของบุคคลที่มีสุขภาพดี (A) และผู้ป่วย (B)
ข้าว. 1.36 อิเล็กโตรโฟรีซิสบนกระดาษของโปรตีนในพลาสมาในเลือดของบุคคลที่มีสุขภาพดี (A) และผู้ป่วย (B)
ฉัน - γ-โกลบูลิน; II - β-โกลบูลิน; สาม -α 2 - โกลบูลิน; IV-α 2 - โกลบูลิน; วี - อัลบูมิน
หลายโรคมาพร้อมกับการเปลี่ยนแปลงเชิงปริมาณในองค์ประกอบของเวย์โปรตีน (dysproteinemia) ลักษณะของการเปลี่ยนแปลงเหล่านี้จะถูกนำมาพิจารณาเมื่อทำการวินิจฉัยและประเมินความรุนแรงและระยะของโรค
โดยใช้ข้อมูลที่ให้ไว้ในตาราง 1.5 ตั้งสมมติฐานเกี่ยวกับโรคซึ่งมีลักษณะโดยโปรไฟล์อิเล็กโตรโฟรีติกที่แสดงในรูปที่ 1.36.
ตารางที่ 1.5. การเปลี่ยนแปลงความเข้มข้นของโปรตีนในเลือดในเลือดในทางพยาธิวิทยา