นักชีววิทยาระดับโมเลกุล อณูชีววิทยาประยุกต์
อณูชีววิทยาประสบกับช่วงเวลาของการพัฒนาอย่างรวดเร็วของวิธีการวิจัยของตนเอง ซึ่งตอนนี้แตกต่างจากชีวเคมี ซึ่งรวมถึงโดยเฉพาะอย่างยิ่ง วิธีการทางพันธุวิศวกรรม การโคลนนิ่ง การแสดงออกทางประดิษฐ์ และการทำให้ล้มลงของยีน เนื่องจาก DNA เป็นสื่อนำพาข้อมูลทางพันธุศาสตร์ ชีววิทยาระดับโมเลกุลจึงใกล้ชิดกับพันธุศาสตร์มากขึ้น และอณูพันธุศาสตร์ก็ก่อตัวขึ้นที่ทางแยก ซึ่งเป็นทั้งส่วนหนึ่งของพันธุศาสตร์และอณูชีววิทยา เช่นเดียวกับที่อณูชีววิทยาใช้ไวรัสเป็นเครื่องมือในการวิจัยอย่างกว้างขวาง ไวรัสวิทยาใช้วิธีการของอณูชีววิทยาเพื่อแก้ปัญหา เทคโนโลยีคอมพิวเตอร์มีส่วนร่วมในการวิเคราะห์ข้อมูลทางพันธุกรรมที่เกี่ยวข้องกับพื้นที่ใหม่ของพันธุศาสตร์ระดับโมเลกุลซึ่งบางครั้งถือว่าเป็นสาขาวิชาพิเศษ: ชีวสารสนเทศ, จีโนมและโปรตีโอมิกส์
ประวัติการพัฒนา
การค้นพบนี้จัดทำขึ้นโดยการวิจัยระยะยาวนานเกี่ยวกับพันธุศาสตร์และชีวเคมีของไวรัสและแบคทีเรีย
ในปีพ.ศ. 2471 เฟรเดอริก กริฟฟิธได้แสดงให้เห็นเป็นครั้งแรกว่าสารสกัดจากแบคทีเรียก่อโรคที่ฆ่าด้วยความร้อนสามารถถ่ายทอดลักษณะการก่อโรคไปสู่แบคทีเรียที่ไม่เป็นพิษเป็นภัย การศึกษาการเปลี่ยนแปลงของแบคทีเรียนำไปสู่การทำให้บริสุทธิ์ของโรคซึ่งตรงกันข้ามกับความคาดหวังกลับกลายเป็นไม่ใช่โปรตีน แต่เป็นกรดนิวคลีอิก กรดนิวคลีอิกไม่เป็นอันตราย แต่มียีนที่กำหนดการก่อโรคและคุณสมบัติอื่นๆ ของจุลินทรีย์เท่านั้น
ในยุค 50 ของศตวรรษที่ XX แสดงให้เห็นว่าแบคทีเรียมีกระบวนการทางเพศแบบดั้งเดิม พวกเขาสามารถแลกเปลี่ยน DNA นอกโครโมโซม, พลาสมิดได้ การค้นพบพลาสมิด เช่นเดียวกับการเปลี่ยนแปลง ทำให้เกิดพื้นฐานของเทคโนโลยีพลาสมิดที่พบได้ทั่วไปในอณูชีววิทยา การค้นพบที่สำคัญอีกประการหนึ่งสำหรับวิธีการนี้คือการค้นพบไวรัสแบคทีเรีย bacteriophages เมื่อต้นศตวรรษที่ 20 ฟาจยังสามารถถ่ายโอนสารพันธุกรรมจากเซลล์แบคทีเรียหนึ่งไปยังอีกเซลล์หนึ่ง การติดเชื้อแบคทีเรียโดยฟาจทำให้เกิดการเปลี่ยนแปลงในองค์ประกอบของอาร์เอ็นเอของแบคทีเรีย หากปราศจากฟาจ องค์ประกอบของ RNA จะคล้ายกับองค์ประกอบของ DNA ของแบคทีเรีย หลังจากการติดเชื้อ RNA จะคล้ายกับ DNA ของแบคทีเรียมากขึ้น ดังนั้นจึงพบว่าโครงสร้างของ RNA ถูกกำหนดโดยโครงสร้างของ DNA ในทางกลับกัน อัตราการสังเคราะห์โปรตีนในเซลล์ขึ้นอยู่กับปริมาณของสารเชิงซ้อน RNA-โปรตีน ได้สูตรมาตามนี้ค่ะ หลักความเชื่อของอณูชีววิทยา:ดีเอ็นเอ ↔ RNA → โปรตีน
การพัฒนาต่อไปของอณูชีววิทยามาพร้อมกับทั้งการพัฒนาวิธีการของมัน โดยเฉพาะอย่างยิ่ง การประดิษฐ์วิธีการในการกำหนดลำดับนิวคลีโอไทด์ของ DNA (W. Gilbert และ F. Sanger, รางวัลโนเบลสาขาเคมีในปี 1980) และใหม่ การค้นพบในด้านการวิจัยเกี่ยวกับโครงสร้างและการทำงานของยีน (ดู ประวัติพันธุกรรม) เมื่อต้นศตวรรษที่ 21 ได้รับข้อมูลเกี่ยวกับโครงสร้างหลักของ DNA ของมนุษย์ทั้งหมดและสิ่งมีชีวิตอื่นๆ อีกจำนวนหนึ่ง ซึ่งมีความสำคัญมากที่สุดสำหรับการแพทย์ การเกษตร และการวิจัยทางวิทยาศาสตร์ ซึ่งนำไปสู่การเกิดขึ้นของพื้นที่ใหม่หลายด้านในชีววิทยา: จีโนมิกส์ , ชีวสารสนเทศ เป็นต้น
ดูสิ่งนี้ด้วย
- อณูชีววิทยา (วารสาร)
- ทรานสคริปโทมิกส์
- ซากดึกดำบรรพ์ระดับโมเลกุล
- EMBO - องค์การยุโรปเพื่ออณูชีววิทยา
วรรณกรรม
- นักร้อง M. , Berg P.ยีนและจีโนม - มอสโก, 1998.
- Stent G. , Kalindar R.พันธุศาสตร์ระดับโมเลกุล - มอสโก, 1981.
- Sambrook J. , Fritsch E.F. , Maniatis T.การโคลนโมเลกุล - 1989.
- Patrushev L.I.การแสดงออกของยีน - ม.: เนาก้า, 2000. - 000 p., ป่วย. ISBN 5-02-001890-2
ลิงค์
มูลนิธิวิกิมีเดีย 2010 .
- เขต Ardatovsky ของภูมิภาค Nizhny Novgorod
- เขต Arzamas ของภูมิภาค Nizhny Novgorod
ดูว่า "อณูชีววิทยา" ในพจนานุกรมอื่นๆ คืออะไร:
อณูชีววิทยา- ศึกษาพื้นฐาน คุณสมบัติและอาการแสดงของสิ่งมีชีวิตในระดับโมเลกุล ทิศทางที่สำคัญที่สุดใน M. b. เป็นการศึกษาโครงสร้างและหน้าที่ของเครื่องมือทางพันธุกรรมของเซลล์และกลไกในการดำเนินการข้อมูลทางพันธุกรรม ... ... พจนานุกรมสารานุกรมชีวภาพ
อณูชีววิทยา- สำรวจคุณสมบัติพื้นฐานและอาการแสดงของชีวิตในระดับโมเลกุล ค้นพบวิธีการและขอบเขตการเจริญเติบโตและการพัฒนาของสิ่งมีชีวิต การจัดเก็บและการส่งข้อมูลทางพันธุกรรม การแปลงพลังงานในเซลล์ของสิ่งมีชีวิต และปรากฏการณ์อื่น ๆ อันเนื่องมาจาก ... พจนานุกรมสารานุกรมขนาดใหญ่
อณูชีววิทยา สารานุกรมสมัยใหม่
อณูชีววิทยา- MOLECULAR BIOLOGY การศึกษาทางชีววิทยาของโครงสร้างและหน้าที่ของ MOLECULES ที่ประกอบเป็นสิ่งมีชีวิต สาขาวิชาหลักของการศึกษาประกอบด้วยคุณสมบัติทางกายภาพและทางเคมีของโปรตีนและกรดนิวคลีอิก เช่น ดีเอ็นเอ ดูสิ่งนี้ด้วย… … พจนานุกรมสารานุกรมวิทยาศาสตร์และเทคนิค
อณูชีววิทยา- ส่วนหนึ่งของ biol. ซึ่งสำรวจคุณสมบัติพื้นฐานและอาการแสดงของชีวิตในระดับโมเลกุล. ค้นพบวิธีการและขอบเขตการเจริญเติบโตและการพัฒนาของสิ่งมีชีวิต การจัดเก็บและการส่งข้อมูลทางพันธุกรรม การแปลงพลังงานในเซลล์ของสิ่งมีชีวิตและ ... ... พจนานุกรมจุลชีววิทยา
อณูชีววิทยา- — หัวข้อเทคโนโลยีชีวภาพ EN ชีววิทยาระดับโมเลกุล … คู่มือนักแปลทางเทคนิค
อณูชีววิทยา- MOLECULAR BIOLOGY สำรวจคุณสมบัติพื้นฐานและอาการแสดงของชีวิตในระดับโมเลกุล ค้นพบวิธีการและขอบเขตการเจริญเติบโตและการพัฒนาของสิ่งมีชีวิต การจัดเก็บและการส่งข้อมูลทางพันธุกรรม การแปลงพลังงานในเซลล์ของสิ่งมีชีวิตและ ... ... พจนานุกรมสารานุกรมภาพประกอบ
อณูชีววิทยา- วิทยาศาสตร์ที่กำหนดเป็นหน้าที่ของความรู้เกี่ยวกับธรรมชาติของปรากฏการณ์ชีวิตโดยการศึกษาวัตถุและระบบทางชีววิทยาในระดับที่เข้าใกล้ระดับโมเลกุลและในบางกรณีถึงขีดจำกัดนี้. เป้าหมายสุดท้ายของเรื่องนี้คือ…… สารานุกรมแห่งสหภาพโซเวียตผู้ยิ่งใหญ่
อณูชีววิทยา- ศึกษาปรากฏการณ์ของชีวิตในระดับโมเลกุลขนาดใหญ่ (ch. arr. โปรตีนและกรดนิวคลีอิก) ในโครงสร้างที่ปราศจากเซลล์ (ไรโบโซม ฯลฯ) ในไวรัสและในเซลล์ด้วย วัตถุประสงค์ของม. การสร้างบทบาทและกลไกการทำงานของโมเลกุลขนาดใหญ่เหล่านี้ตาม ... ... สารานุกรมเคมี
อณูชีววิทยา- สำรวจคุณสมบัติพื้นฐานและอาการแสดงของชีวิตในระดับโมเลกุล ค้นพบวิธีการและขอบเขตการเจริญเติบโตและการพัฒนาของสิ่งมีชีวิตการจัดเก็บและการส่งข้อมูลทางพันธุกรรมการแปลงพลังงานในเซลล์ของสิ่งมีชีวิตและปรากฏการณ์อื่น ๆ ... ... พจนานุกรมสารานุกรม
หนังสือ
- อณูชีววิทยาของเซลล์ หนังสือปัญหา, เจ. วิลสัน, ที. ฮันท์ หนังสือของนักประพันธ์ชาวอเมริกันเป็นภาคผนวกของหนังสือเรียน 'Molecular Biology of the Cell' ฉบับที่ 2 โดย B. Alberts, D. Bray, J. Lewis และคนอื่นๆ มีคำถามและงานซึ่งมีจุดประสงค์เพื่อทำให้ลึกซึ้งยิ่งขึ้น ..
อณูชีววิทยาวิทยาศาสตร์ที่กำหนดเป็นหน้าที่ของความรู้เกี่ยวกับธรรมชาติของปรากฏการณ์ชีวิตโดยการศึกษาวัตถุและระบบทางชีววิทยาในระดับที่เข้าใกล้ระดับโมเลกุลและในบางกรณีถึงขีดจำกัดนี้ เป้าหมายสูงสุดในกรณีนี้คือการค้นหาวิธีการและขอบเขตของการสำแดงลักษณะเฉพาะของชีวิต เช่น การถ่ายทอดทางพันธุกรรม การสืบพันธุ์ในแบบของตัวเอง การสังเคราะห์โปรตีน ความตื่นเต้นง่าย การเติบโตและการพัฒนา การจัดเก็บและการส่งข้อมูล การแปลงพลังงาน การเคลื่อนที่ เป็นต้น เกิดจากโครงสร้าง คุณสมบัติ และปฏิสัมพันธ์ของโมเลกุลของสารที่มีความสำคัญทางชีววิทยา โดยหลักแล้วคือ โปรตีนและกรดนิวคลีอิกสองกลุ่มหลักหลัก ลักษณะเด่นของเอ็มบี - การศึกษาปรากฏการณ์แห่งชีวิตบนวัตถุที่ไม่มีชีวิตหรือสิ่งที่มีลักษณะเฉพาะโดยอาการดึกดำบรรพ์ที่สุดของชีวิต นี่คือการก่อตัวทางชีวภาพตั้งแต่ระดับเซลล์และต่ำกว่า: ออร์แกเนลล์ย่อย เช่น นิวเคลียสของเซลล์ที่แยกได้ ไมโทคอนเดรีย ไรโบโซม โครโมโซม เยื่อหุ้มเซลล์ เพิ่มเติม - ระบบที่ยืนอยู่บนพรมแดนของธรรมชาติที่มีชีวิตและไม่มีชีวิต - ไวรัสรวมถึงแบคทีเรียและจบลงด้วยโมเลกุลของส่วนประกอบที่สำคัญที่สุดของสิ่งมีชีวิต - กรดนิวคลีอิกและโปรตีน
รากฐานที่เอ็มพัฒนาขึ้น มีการเชื่อมโยงอย่างแยกไม่ออกกับอณูพันธุศาสตร์ ซึ่งยังคงเป็นส่วนสำคัญของ
ลักษณะเด่นของเอ็มบี เป็นสามมิติของมัน สาระสำคัญของ M. b. M. Perutz มองเห็นในการตีความหน้าที่ทางชีววิทยาในแง่ของโครงสร้างโมเลกุล เอ็มบี มีวัตถุประสงค์เพื่อให้ได้คำตอบสำหรับคำถาม "อย่างไร" รู้สาระสำคัญของบทบาทและการมีส่วนร่วมของโครงสร้างทั้งหมดของโมเลกุลและสำหรับคำถาม "ทำไม" และ "เพื่ออะไร" ค้นหาความสัมพันธ์ในด้านหนึ่ง ระหว่างคุณสมบัติของโมเลกุล (เช่น โปรตีนและกรดนิวคลีอิกเป็นหลัก) และหน้าที่ของมัน และในทางกลับกัน บทบาทของหน้าที่ส่วนบุคคลดังกล่าวในความซับซ้อนโดยรวมของการแสดงอาการของกิจกรรมที่สำคัญ
ความสำเร็จที่สำคัญที่สุดของอณูชีววิทยายังห่างไกลจากรายชื่อความสำเร็จทั้งหมด: การเปิดเผยโครงสร้างและกลไกการทำงานทางชีววิทยาของ DNA, RNA และไรโบโซมทุกประเภท, การเปิดเผยรหัสพันธุกรรม; การค้นพบการถอดรหัสแบบย้อนกลับ กล่าวคือ การสังเคราะห์ DNA บนเทมเพลต RNA การศึกษากลไกการทำงานของเม็ดสีระบบทางเดินหายใจ การค้นพบโครงสร้างสามมิติและบทบาทหน้าที่ของมันในการทำงานของเอนไซม์ หลักการของการสังเคราะห์เมทริกซ์และกลไกของการสังเคราะห์โปรตีน การเปิดเผยโครงสร้างของไวรัสและกลไกของการจำลองแบบ โครงสร้างหลักและบางส่วน โครงสร้างเชิงพื้นที่ของแอนติบอดี การแยกยีนแต่ละตัว การสังเคราะห์ยีนทางเคมีและการสังเคราะห์ทางชีวภาพ (เอนไซม์) รวมทั้งมนุษย์ นอกเซลล์ (ในหลอดทดลอง); การถ่ายโอนยีนจากสิ่งมีชีวิตหนึ่งไปยังอีกสิ่งมีชีวิตหนึ่งรวมถึงไปยังเซลล์ของมนุษย์ การถอดรหัสโครงสร้างทางเคมีที่ก้าวหน้าอย่างรวดเร็วของโปรตีนแต่ละตัวที่เพิ่มขึ้นซึ่งส่วนใหญ่เป็นเอนไซม์และกรดนิวคลีอิก การค้นพบปรากฏการณ์ของ "การประกอบตัวเอง" ของวัตถุทางชีววิทยาบางอย่างที่มีความซับซ้อนเพิ่มขึ้นเรื่อย ๆ โดยเริ่มจากโมเลกุลของกรดนิวคลีอิกและไปสู่เอ็นไซม์หลายองค์ประกอบ ไวรัส ไรโบโซม ฯลฯ การชี้แจงของ allosteric และหลักการพื้นฐานอื่น ๆ ของการควบคุมการทำงานและกระบวนการทางชีววิทยา
ปัญหาทางอณูชีววิทยาพร้อมกับกำหนดภารกิจสำคัญที่เอ็มจะ (ความรู้เกี่ยวกับกฎของ "การรับรู้" การรวมตัวเองและการบูรณาการ) ทิศทางที่แท้จริงของการค้นหาทางวิทยาศาสตร์สำหรับอนาคตอันใกล้คือการพัฒนาวิธีการที่อนุญาตให้ถอดรหัสโครงสร้างแล้วองค์กรสามมิติเชิงพื้นที่ของโมเลกุลสูง กรดนิวคลีอิก. วิธีการที่สำคัญที่สุดทั้งหมดที่ใช้ทำให้แน่ใจถึงการเกิดขึ้นและความสำเร็จของ M b. ได้รับการเสนอและพัฒนาโดยนักฟิสิกส์ (ultracentrifugation การวิเคราะห์การเลี้ยวเบนของรังสีเอกซ์ กล้องจุลทรรศน์อิเล็กตรอน แนวทางการทดลองทางกายภาพแบบใหม่เกือบทั้งหมด (เช่น การใช้คอมพิวเตอร์ ซินโครตรอน หรือเบรมสตราลุง การแผ่รังสี เทคโนโลยีเลเซอร์ และอื่นๆ) เปิดโอกาสให้มีการศึกษาเชิงลึกเกี่ยวกับปัญหาของการวิเคราะห์อุตุนิยมวิทยา ในบรรดาภารกิจที่สำคัญที่สุดในลักษณะที่ใช้งานได้จริง คำตอบที่คาดหวังจาก M. b. ในตอนแรกคือปัญหาของพื้นฐานระดับโมเลกุลของการเติบโตของมะเร็ง จากนั้น - วิธีป้องกันและอาจเอาชนะโรคทางพันธุกรรม - " โรคระดับโมเลกุล" สิ่งที่สำคัญอย่างยิ่งคือการอธิบายพื้นฐานระดับโมเลกุลของการเร่งปฏิกิริยาทางชีววิทยา กล่าวคือ การทำงานของเอนไซม์ ท่ามกลางทิศทางสมัยใหม่ที่สำคัญที่สุดของ M. b. ควรรวมถึงความปรารถนาที่จะถอดรหัสกลไกระดับโมเลกุลของการกระทำของฮอร์โมนสารพิษและยารวมทั้งค้นหารายละเอียดของโครงสร้างโมเลกุลและการทำงานของโครงสร้างเซลล์เช่นเยื่อหุ้มชีวภาพที่เกี่ยวข้องกับการควบคุมกระบวนการแทรกซึมและ การขนส่งสาร เป้าหมายที่ห่างไกลมากขึ้น M. b. - ความรู้เกี่ยวกับธรรมชาติของกระบวนการทางประสาท กลไกของความจำ ฯลฯ หนึ่งในส่วนที่เกิดขึ้นที่สำคัญของ M. b. - ที่เรียกว่า. พันธุวิศวกรรมซึ่งมีจุดมุ่งหมายเพื่อดำเนินการเครื่องมือทางพันธุกรรม (จีโนม) ของสิ่งมีชีวิตโดยเจตนาโดยเริ่มจากจุลินทรีย์และต่ำกว่า (เซลล์เดียว) และสิ้นสุดที่มนุษย์ (ในกรณีหลังส่วนใหญ่เพื่อวัตถุประสงค์ในการรักษาโรคทางพันธุกรรมอย่างรุนแรงและการแก้ไข ความบกพร่องทางพันธุกรรม)
ทิศทางที่สำคัญที่สุดของ MB:
- พันธุศาสตร์ระดับโมเลกุล - การศึกษาโครงสร้างและการทำงานของโครงสร้างและการทำงานของเครื่องมือทางพันธุกรรมของเซลล์และกลไกในการดำเนินการข้อมูลทางพันธุกรรม
– ไวรัสวิทยาระดับโมเลกุล – การศึกษากลไกระดับโมเลกุลของปฏิสัมพันธ์ของไวรัสกับเซลล์
– ภูมิคุ้มกันวิทยาระดับโมเลกุล – การศึกษารูปแบบของปฏิกิริยาภูมิคุ้มกันของร่างกาย
– อณูชีววิทยาของการพัฒนา – การศึกษาการปรากฏตัวของความหลากหลายของเซลล์ในระหว่างการพัฒนาบุคคลของสิ่งมีชีวิตและความเชี่ยวชาญของเซลล์
วัตถุประสงค์หลักของการวิจัย: ไวรัส (รวมถึงแบคทีเรีย) เซลล์และโครงสร้างย่อยของเซลล์ โมเลกุลขนาดใหญ่ สิ่งมีชีวิตหลายเซลล์
การพัฒนาทางชีวเคมี ชีวฟิสิกส์ พันธุศาสตร์ ไซโตเคมี จุลชีววิทยาและไวรัสวิทยาหลายส่วนในช่วงต้นทศวรรษที่ 40 ของศตวรรษที่ XX นำไปสู่การศึกษาปรากฏการณ์ชีวิตในระดับโมเลกุลอย่างใกล้ชิด ความสำเร็จที่สำเร็จโดยวิทยาศาสตร์เหล่านี้พร้อมกันและจากด้านต่าง ๆ นำไปสู่ความจริงที่ว่ามันอยู่ในระดับโมเลกุลที่ระบบควบคุมหลักของการทำงานของร่างกายและความก้าวหน้าต่อไปของวิทยาศาสตร์เหล่านี้จะขึ้นอยู่กับการเปิดเผยของ หน้าที่ทางชีววิทยาของโมเลกุลที่ประกอบเป็นร่างกายของสิ่งมีชีวิต การมีส่วนร่วมในการสังเคราะห์และการสลายตัว การเปลี่ยนแปลงร่วมกันและการสืบพันธุ์ของสารประกอบในเซลล์ ตลอดจนการแลกเปลี่ยนพลังงานและข้อมูลที่เกิดขึ้นในกรณีนี้ ดังนั้นที่จุดเชื่อมต่อของสาขาวิชาทางชีววิทยาเหล่านี้กับเคมีและฟิสิกส์สาขาใหม่อย่างสมบูรณ์ - ชีววิทยาระดับโมเลกุล
แตกต่างจากชีวเคมี ความสนใจของอณูชีววิทยาสมัยใหม่มุ่งเน้นไปที่การศึกษาโครงสร้างและหน้าที่ของคลาสที่สำคัญที่สุดของไบโอพอลิเมอร์ - โปรตีนและกรดนิวคลีอิกซึ่งในตอนแรกกำหนดความเป็นไปได้ของปฏิกิริยาเมแทบอลิซึมและประการที่สอง - การสังเคราะห์โปรตีนจำเพาะ ดังนั้นจึงเป็นที่ชัดเจนว่า เป็นไปไม่ได้ที่จะสร้างความแตกต่างที่ชัดเจนระหว่างอณูชีววิทยาและชีวเคมี ซึ่งเป็นสาขาที่เกี่ยวข้องกันของพันธุศาสตร์ จุลชีววิทยา และไวรัสวิทยา
การเกิดขึ้นของอณูชีววิทยามีความสัมพันธ์อย่างใกล้ชิดกับการพัฒนาวิธีการวิจัยใหม่ ซึ่งได้มีการกล่าวถึงไปแล้วในบทที่เกี่ยวข้อง นอกเหนือจากการพัฒนากล้องจุลทรรศน์อิเล็กตรอนและวิธีการอื่น ๆ ของเทคนิคด้วยกล้องจุลทรรศน์แล้ว วิธีการแยกส่วนขององค์ประกอบเซลล์ที่พัฒนาขึ้นในปี 1950 มีบทบาทสำคัญ โดยอาศัยวิธีการหมุนเหวี่ยงแบบดิฟเฟอเรนเชียลที่ได้รับการปรับปรุง (A. Claude, 1954) ถึงเวลานี้ มีวิธีการที่เชื่อถือได้อยู่แล้วสำหรับการแยกและการแยกส่วนของพอลิเมอร์ชีวภาพ ซึ่งรวมถึงโดยเฉพาะอย่างยิ่ง วิธีการแยกโปรตีนด้วยอิเล็กโตรโฟรีซิสที่เสนอโดย A. Tiselius (1937; Nobel Prize, 1948) วิธีการแยกและทำให้กรดนิวคลีอิกบริสุทธิ์ (E. Kay, A. Downs, M. Sevag, A. Mirsky และอื่นๆ) ในเวลาเดียวกัน วิธีต่างๆ ของการวิเคราะห์ด้วยโครมาโตกราฟีได้รับการพัฒนาในห้องปฏิบัติการหลายแห่งของโลก (A. Martin and R. Sing, 1941; Nobel Prize, 1952) ต่อมาได้รับการปรับปรุงอย่างมีนัยสำคัญ
การวิเคราะห์การเลี้ยวเบนของรังสีเอกซ์ถือเป็นบริการที่ประเมินค่าไม่ได้ในการถอดรหัสโครงสร้างของไบโอโพลีเมอร์ หลักการพื้นฐานของการวิเคราะห์การเลี้ยวเบนของรังสีเอกซ์ได้รับการพัฒนาที่ King's College London University ภายใต้การนำของ W. Bragg โดยกลุ่มนักวิจัย ซึ่งรวมถึง J. Bernal, A. Londsdale, W. Astbury, J. Robertson และคนอื่นๆ
ควรกล่าวถึงเป็นพิเศษเกี่ยวกับการศึกษา Protoplasm Biochemistry (1925 - 1929) ศาสตราจารย์แห่งมหาวิทยาลัยแห่งรัฐมอสโก A.R. Kizel ซึ่งมีความสำคัญอย่างยิ่งต่อการพัฒนาชีววิทยาระดับโมเลกุลในภายหลัง Kizel จัดการกับความคิดที่หยั่งรากอย่างแน่นหนาว่าโปรโตพลาสซึมใด ๆ ขึ้นอยู่กับโปรตีนในร่างกาย - แผ่นซึ่งถูกกล่าวหาว่ากำหนดคุณสมบัติโครงสร้างและการทำงานที่สำคัญที่สุดทั้งหมด เขาแสดงให้เห็นว่าแผ่นเปลือกโลกเป็นโปรตีนที่พบเฉพาะในไมโซไมซีเตส และจากนั้นก็อยู่ในระยะหนึ่งของการพัฒนา และไม่มีส่วนประกอบถาวร - โปรตีนโครงกระดูกตัวเดียว - อยู่ในโปรโตพลาสซึม ดังนั้นการศึกษาปัญหาโครงสร้างของโปรโตพลาสซึมและบทบาทหน้าที่ของโปรตีนจึงเป็นแนวทางที่ถูกต้องและได้รับขอบเขตสำหรับการพัฒนา การวิจัยของ Kisel ได้รับการยอมรับจากทั่วโลก โดยกระตุ้นการศึกษาเคมีของส่วนประกอบต่างๆ ของเซลล์
คำว่า "อณูชีววิทยา" ซึ่งใช้ครั้งแรกโดยศาสตราจารย์ด้านผลึกศาสตร์ชาวอังกฤษแห่งมหาวิทยาลัยลีดส์ ดับเบิลยู. แอสท์บิวรี อาจปรากฏขึ้นในช่วงต้นทศวรรษ 1940 (ก่อนปี ค.ศ. 1945) การศึกษาการเลี้ยวเบนของรังสีเอกซ์พื้นฐานของโปรตีนและ DNA ที่ดำเนินการโดย Astbury ในช่วงทศวรรษที่ 1930 นั้นเป็นพื้นฐานสำหรับการถอดรหัสโครงสร้างรองของไบโอโพลีเมอร์เหล่านี้ที่ประสบความสำเร็จในภายหลัง ในปี 1963 J. Bernal เขียนว่า: "อนุสาวรีย์สำหรับเขาจะถูกสร้างโดยชีววิทยาโมเลกุลทั้งหมด - วิทยาศาสตร์ที่เขาตั้งชื่อและก่อตั้งจริงๆ" * , ในวรรณคดีคำนี้ปรากฏขึ้นเป็นครั้งแรกบางทีในปี 1946 ในบทความโดย W. Astbury "ความคืบหน้าในการวิเคราะห์การเลี้ยวเบนของรังสีเอกซ์ของสารประกอบอินทรีย์และไฟบริลลาร์" ซึ่งตีพิมพ์ในวารสารภาษาอังกฤษ "Nature" ** ในการบรรยาย Harvey Lecture ของเขา Astbury (1950) กล่าวว่า “ฉันยินดีที่ตอนนี้มีการใช้คำว่าอณูชีววิทยาอย่างแพร่หลาย แม้ว่าไม่น่าจะเป็นไปได้ที่ฉันจะเป็นคนแรกที่เสนอ ฉันชอบมันและฉันพยายามจะเผยแพร่มานานแล้ว ” ***. ในปี 1950 Astbury นั้นชัดเจนว่าอณูชีววิทยาเกี่ยวข้องกับโครงสร้างและโครงสร้างของโมเลกุลขนาดใหญ่เป็นหลัก การศึกษาซึ่งมีความสำคัญอย่างยิ่งต่อการทำความเข้าใจการทำงานของสิ่งมีชีวิต
* (ไบโอจี เมม. เพื่อนรอย. Soc, 1963, วี. 9, 29.)
** (ดับเบิลยู. ที. แอสท์บิวรี. ความคืบหน้าของการวิเคราะห์ X-ray ของโครงสร้างอินทรีย์และเส้นใย.- ธรรมชาติ,. 2489 วี. 157, 121.)
*** (ดับเบิลยู. ที. แอสท์บิวรี. การผจญภัยในอณูชีววิทยา. โธมัส สปริงฟิลด์, 1952, p. 3.)
อณูชีววิทยาเผชิญและเผชิญในความเป็นจริงงานเดียวกันกับชีววิทยาโดยรวม - ความรู้เกี่ยวกับสาระสำคัญของชีวิตและปรากฏการณ์พื้นฐานโดยเฉพาะอย่างยิ่งเช่นการถ่ายทอดทางพันธุกรรมและความแปรปรวน อณูชีววิทยาสมัยใหม่มีวัตถุประสงค์หลักเพื่อถอดรหัสโครงสร้างและหน้าที่ของยีน วิธีการและกลไกในการทำความเข้าใจข้อมูลทางพันธุกรรมของสิ่งมีชีวิตในขั้นตอนต่างๆ ของการสร้างยีนและในระยะต่างๆ ของการอ่าน มันถูกออกแบบมาเพื่อเปิดเผยกลไกที่ละเอียดอ่อนของการควบคุมการทำงานของยีนและการสร้างความแตกต่างของเซลล์ เพื่ออธิบายธรรมชาติของการกลายพันธุ์และพื้นฐานระดับโมเลกุลของกระบวนการวิวัฒนาการ
การสร้างบทบาททางพันธุกรรมของกรดนิวคลีอิก
สำหรับการพัฒนาอณูชีววิทยา การค้นพบต่อไปนี้มีความสำคัญมากที่สุด ในปี 1944 นักวิจัยชาวอเมริกัน O. Avery, K. McLeod (รางวัลโนเบล, 1923) และ M. McCarthy แสดงให้เห็นว่าโมเลกุลของ DNA ที่แยกได้จาก pneumococci ได้เปลี่ยนแปลงกิจกรรม หลังจากการไฮโดรไลซิสของ DNA เหล่านี้โดย deoxyribonuclease กิจกรรมการเปลี่ยนแปลงของพวกมันก็หายไปอย่างสมบูรณ์ ด้วยเหตุนี้ จึงได้รับการพิสูจน์อย่างน่าเชื่อถือเป็นครั้งแรกว่านี่คือ DNA ไม่ใช่โปรตีน ที่มีหน้าที่ทางพันธุกรรมในเซลล์
เพื่อความเป็นธรรม ควรสังเกตว่าปรากฏการณ์ของการเปลี่ยนแปลงของแบคทีเรียถูกค้นพบเร็วกว่าการค้นพบ Avery, McLeod และ McCarthy ในปีพ.ศ. 2471 เอฟ. กริฟฟิธได้ตีพิมพ์บทความซึ่งเขารายงานว่าหลังจากเพิ่มเซลล์ที่ฆ่าแล้วของสายพันธุ์ที่มีเชื้อก่อโรคที่ห่อหุ้มไปยังปอดบวมที่ไม่ก่อให้เกิดโรค (ไม่ห่อหุ้ม) ส่วนผสมที่เป็นผลลัพธ์ของเซลล์จะกลายเป็นอันตรายถึงชีวิตสำหรับหนูทดลอง นอกจากนี้ เซลล์นิวโมคอคคัสที่มีชีวิตซึ่งแยกได้จากสัตว์ที่ติดเชื้อด้วยส่วนผสมนี้มีความรุนแรงอยู่แล้วและมีแคปซูลโพลีแซ็กคาไรด์ ดังนั้น ในการทดลองนี้ แสดงให้เห็นว่าภายใต้อิทธิพลของส่วนประกอบบางอย่างของเซลล์นิวโมคอคคัสที่ถูกฆ่า รูปแบบของแบคทีเรียที่ไม่ห่อหุ้มจะเปลี่ยนเป็นรูปแบบที่ก่อโรคร้ายที่ก่อตัวเป็นแคปซูล สิบหกปีต่อมา Avery, McLeod และ McCarthy ได้แทนที่เซลล์นิวโมคอคคัสที่ฆ่าแล้วทั้งหมดด้วยกรดดีออกซีไรโบนิวคลีอิกในการทดลองนี้ และแสดงให้เห็นว่าเป็น DNA ที่มีการเปลี่ยนแปลงกิจกรรม (ดูบทที่ 7 และ 25 ด้วย) ความสำคัญของการค้นพบนี้เป็นเรื่องยากที่จะประเมินค่าสูงไป โดยกระตุ้นการศึกษากรดนิวคลีอิกในห้องปฏิบัติการหลายแห่งทั่วโลก และบังคับให้นักวิทยาศาสตร์ให้ความสำคัญกับดีเอ็นเอ
ควบคู่ไปกับการค้นพบเอเวอรี แมคเลียด และแม็กคาร์ธี เมื่อต้นทศวรรษ 1950 มีหลักฐานทางตรงและทางอ้อมจำนวนมากพอสมควรได้สะสมไว้แล้วว่ากรดนิวคลีอิกมีบทบาทสำคัญในชีวิตและมีหน้าที่ทางพันธุกรรม โดยเฉพาะอย่างยิ่งสิ่งนี้ถูกระบุโดยธรรมชาติของการแปล DNA ในเซลล์และข้อมูลของ R. Vendrelli (1948) ที่เนื้อหา DNA ต่อเซลล์นั้นคงที่อย่างเคร่งครัดและสัมพันธ์กับระดับของ ploidy: ในเซลล์สืบพันธุ์เดี่ยว DNA คือ ครึ่งหนึ่งในเซลล์โซมาติกแบบดิพลอยด์ ความเสถียรทางเมตาบอลิซึมที่เด่นชัดของ DNA ยังพิสูจน์ให้เห็นถึงบทบาททางพันธุกรรมของ DNA ในตอนต้นของยุค 50 มีข้อเท็จจริงหลายอย่างสะสมอยู่ ซึ่งบ่งชี้ว่าปัจจัยการกลายพันธุ์ที่รู้จักส่วนใหญ่ทำหน้าที่หลักในกรดนิวคลีอิก และโดยเฉพาะอย่างยิ่งใน DNA (R. Hotchkiss, 1949; G. Ephrussi-Taylor, 1951; E. Freese , 2500 และอื่น ๆ).
สิ่งสำคัญอย่างยิ่งในการสร้างบทบาททางพันธุกรรมของกรดนิวคลีอิกคือการศึกษาฟาจและไวรัสต่างๆ ในปีพ.ศ. 2476 ดี. ชเลซิงเงอร์พบดีเอ็นเอในแบคทีเรียในแบคทีเรีย Escherichia coli นับตั้งแต่การแยกไวรัสโมเสคจากยาสูบ (TMV) ในสถานะผลึกโดยดับเบิลยู สแตนลีย์ (1935, รางวัลโนเบล, 2489) เวทีใหม่ในการศึกษาไวรัสพืชได้เริ่มต้นขึ้น ในปี 2480 - 2481 พนักงานของสถานีเกษตร Rothamsted (อังกฤษ) F. Bowden และ N. Pirie แสดงให้เห็นว่าไวรัสพืชหลายชนิดที่แยกได้จากพวกมันไม่ใช่โกลบูลิน แต่เป็นไรโบนิวคลีโอโปรตีนและมีกรดนิวคลีอิกเป็นส่วนประกอบบังคับ ในตอนต้นของยุค 40 ผลงานของ G. Schramm (1940), P. A. Agatov (1941), G. Miller และ W. Stanley (1941) ได้รับการตีพิมพ์ซึ่งระบุว่าการดัดแปลงทางเคมีที่เห็นได้ชัดเจนของส่วนประกอบโปรตีนไม่ได้นำไปสู่ ต่อการสูญเสียการติดเชื้อ TMV สิ่งนี้บ่งชี้ว่าส่วนประกอบโปรตีนไม่สามารถเป็นพาหะของคุณสมบัติทางพันธุกรรมของไวรัสได้ เนื่องจากนักจุลชีววิทยาหลายคนยังคงเชื่อ หลักฐานที่น่าเชื่อถือสนับสนุนบทบาททางพันธุกรรมของกรดนิวคลีอิก (RNA) ในไวรัสพืชได้รับในปี 1956 โดย G. Schramm ในทูบิงเกน (FRG) และ H. Frenkel-Konrath ในแคลิฟอร์เนีย (สหรัฐอเมริกา) นักวิจัยเหล่านี้เกือบจะพร้อมกันและเป็นอิสระจากอาร์เอ็นเอที่แยกจากกันจาก TMV และแสดงให้เห็นว่าโปรตีนนี้ไม่มีการติดเชื้อ: เป็นผลมาจากการติดเชื้อของพืชยาสูบด้วย RNA นี้ อนุภาคไวรัสปกติได้ถูกสร้างขึ้นและคูณในพวกมัน ซึ่งหมายความว่า RNA มีข้อมูลสำหรับการสังเคราะห์และการประกอบส่วนประกอบไวรัสทั้งหมด รวมทั้งโปรตีนของไวรัส ในปี 1968 I. G. Atabekov ยืนยันว่าโปรตีนมีบทบาทสำคัญในการติดเชื้อของพืช - ธรรมชาติของโปรตีนกำหนดสเปกตรัมของพืชที่เป็นโฮสต์
ในปี 1957 Frenkel-Konrat ได้สร้าง TMV ขึ้นใหม่จากส่วนประกอบที่เป็นส่วนประกอบ - RNA และโปรตีนเป็นครั้งแรก นอกจากอนุภาคปกติแล้ว เขาได้รับ "ลูกผสม" แบบผสม ซึ่ง RNA มาจากสายพันธุ์หนึ่งและโปรตีนจากอีกสายพันธุ์หนึ่ง การถ่ายทอดทางพันธุกรรมของลูกผสมดังกล่าวถูกกำหนดโดย RNA อย่างสมบูรณ์ และลูกหลานของไวรัสนั้นเป็นของสายพันธุ์ที่ RNA ถูกใช้เพื่อให้ได้อนุภาคผสมเริ่มต้น ต่อมาการทดลองของ A. Gierer, G. Schuster และ G. Schramm (1958) และ G. Witman (1960 - 1966) แสดงให้เห็นว่าการดัดแปลงทางเคมีขององค์ประกอบนิวเคลียส TMV ทำให้เกิดการกลายพันธุ์ต่างๆ ของไวรัสนี้
ในปี 1970 D. Baltimore และ G. Temin พบว่าการถ่ายโอนข้อมูลทางพันธุกรรมสามารถเกิดขึ้นได้ไม่เพียงแค่จาก DNA ไปยัง RNA แต่ในทางกลับกัน พวกเขาพบในไวรัสที่ประกอบด้วย RNA ที่ทำให้เกิดมะเร็ง (oncornaviruses) ซึ่งเป็นเอนไซม์พิเศษที่เรียกว่า reverse transcriptase ซึ่งสามารถสังเคราะห์ DNA เสริมบนสายโซ่ RNA การค้นพบครั้งสำคัญนี้ทำให้สามารถเข้าใจกลไกของการแทรกข้อมูลทางพันธุกรรมของไวรัสที่ประกอบด้วย RNA เข้าไปในจีโนมของโฮสต์ และเพื่อดูลักษณะของการกระทำที่ก่อให้เกิดมะเร็งได้อย่างชัดเจน
การค้นพบกรดนิวคลีอิกและการศึกษาคุณสมบัติของกรดเหล่านี้
คำว่ากรดนิวคลีอิกถูกนำมาใช้โดยนักชีวเคมีชาวเยอรมัน R. Altman ในปี 1889 หลังจากที่สารประกอบเหล่านี้ถูกค้นพบในปี 1869 โดยแพทย์ชาวสวิส F. Miescher มิเชอร์สกัดเซลล์หนองด้วยกรดไฮโดรคลอริกเจือจางเป็นเวลาหลายสัปดาห์ และได้รับวัสดุนิวเคลียร์เกือบบริสุทธิ์ในส่วนที่เหลือ เขาถือว่าวัสดุนี้เป็นลักษณะ "สารของนิวเคลียสของเซลล์และเรียกมันว่านิวเคลียส ในแง่ของคุณสมบัติของนิวเคลียส นิวเคลียสแตกต่างจากโปรตีนอย่างรวดเร็ว: เป็นกรดมากขึ้น ไม่มีกำมะถัน แต่มีฟอสฟอรัสอยู่มาก มันคือ ละลายได้ง่ายในด่าง แต่ไม่ละลายในกรดเจือจาง
มิเชอร์ส่งผลการสังเกตของเขาเกี่ยวกับนิวเคลียสให้ F. Goppe-Seyler เพื่อตีพิมพ์ในวารสาร สารที่เขาอธิบายนั้นผิดปกติมาก (ในเวลานั้นมีเพียงเลซิตินเท่านั้นที่รู้จักสารประกอบที่มีฟอสฟอรัสทางชีวภาพทั้งหมด) ซึ่ง Goppe-Seyler ไม่เชื่อการทดลองของ Misher ส่งต้นฉบับคืนให้เขาและสั่งพนักงานของเขา N. Plosh และ N. Lyubavin ให้ไปที่ ตรวจสอบข้อสรุปของเขาเกี่ยวกับเนื้อหาอื่น ๆ งานของ Miescher "เกี่ยวกับองค์ประกอบทางเคมีของเซลล์หนอง" ถูกตีพิมพ์เมื่อสองปีต่อมา (1871) ในเวลาเดียวกัน ผลงานของ Goppe-Seyler และผู้ทำงานร่วมกันของเขาได้รับการตีพิมพ์เกี่ยวกับองค์ประกอบของเซลล์หนอง เม็ดเลือดแดงของนก งู และเซลล์อื่นๆ ในอีกสามปีข้างหน้า นิวเคลียสถูกแยกออกจากเซลล์สัตว์และยีสต์
ในงานของเขา มิเชอร์ตั้งข้อสังเกตว่าการศึกษาอย่างละเอียดของนิวเคลียสที่แตกต่างกันสามารถนำไปสู่การสร้างความแตกต่างระหว่างกัน ดังนั้นจึงคาดการณ์ถึงความคิดเกี่ยวกับความจำเพาะของกรดนิวคลีอิก ขณะศึกษานมปลาแซลมอน มิเชอร์พบว่านิวเคลียสในนมนั้นอยู่ในรูปของเกลือและเกี่ยวข้องกับโปรตีนหลักซึ่งเขาเรียกว่าโปรทามีน
ในปี 1879 A. Kossel เริ่มศึกษานิวเคลียสในห้องปฏิบัติการของ Goppe-Seyler ในปีพ.ศ. 2424 เขาแยกไฮโปแซนทีนออกจากนิวเคลียส แต่ในขณะนั้นเขายังสงสัยที่มาของเบสนี้และเชื่อว่าไฮโปแซนทีนอาจเป็นผลิตภัณฑ์จากการย่อยสลายของโปรตีน ในปี พ.ศ. 2434 คอสเซลได้ค้นพบอะดีนีน กัวนีน กรดฟอสฟอริก และสารอื่นที่มีคุณสมบัติเป็นน้ำตาล สำหรับการวิจัยเกี่ยวกับเคมีของกรดนิวคลีอิก Kossel ได้รับรางวัลโนเบลในปี 1910
ความก้าวหน้าเพิ่มเติมในการถอดรหัสโครงสร้างของกรดนิวคลีอิกนั้นสัมพันธ์กับการวิจัยของพี. เลวินและเพื่อนร่วมงาน (พ.ศ. 2454 - 2477) ในปี 1911 P. Levin และ V. Jacobs ระบุองค์ประกอบคาร์โบไฮเดรตของอะดีโนซีนและกัวโนซีน พวกเขาพบว่านิวคลีโอไซด์เหล่านี้มีดี-ไรโบส ในปีพ.ศ. 2473 เลวินแสดงให้เห็นว่าองค์ประกอบคาร์โบไฮเดรตของดีออกซีไรโบนิวคลีโอไซด์คือ 2-ดีออกซี-ดี-ไรโบส จากผลงานของเขา เป็นที่ทราบกันดีว่ากรดนิวคลีอิกถูกสร้างขึ้นจากนิวคลีโอไทด์ เช่น ฟอสโฟรีเลต นิวคลีโอไซด์ เลวินเชื่อว่าพันธะหลักในกรดนิวคลีอิก (RNA) คือพันธะฟอสโฟไดสเตอร์ 2", 5" ความคิดนี้กลายเป็นสิ่งที่ผิด ต้องขอบคุณผลงานของนักเคมีชาวอังกฤษ A. Todd (รางวัลโนเบล, 2500) และผู้ร่วมงานของเขา เช่นเดียวกับนักชีวเคมีชาวอังกฤษ R. Markham และ J. Smith ในช่วงต้นทศวรรษ 50s เป็นที่ทราบกันดีว่าพันธะประเภทหลักใน RNA คือ 3", 5" - พันธะฟอสโฟไดสเตอร์
Lewin แสดงให้เห็นว่ากรดนิวคลีอิกที่แตกต่างกันสามารถแตกต่างกันในธรรมชาติขององค์ประกอบคาร์โบไฮเดรต: บางส่วนมีน้ำตาลดีออกซีไรโบส ในขณะที่บางชนิดมีไรโบส นอกจากนี้ กรดนิวคลีอิกทั้งสองชนิดนี้ยังมีลักษณะที่แตกต่างกันในธรรมชาติของเบสตัวใดตัวหนึ่ง ได้แก่ กรดนิวคลีอิกประเภทเพนโตสมียูราซิล และกรดนิวคลีอิกประเภทดีออกซีเพนโทสมีไทมีน กรดนิวคลีอิกดีออกซีเพนโทส (ในคำศัพท์สมัยใหม่ กรดดีออกซีไรโบนิวคลีอิก - DNA) มักจะแยกออกได้ง่ายในปริมาณมากจากต่อมไทมัส (ต่อมหวาน) ของน่อง ดังนั้นจึงเรียกว่ากรดไทโมนิวคลีอิก แหล่งที่มาของกรดนิวคลีอิกชนิดเพนโตส (RNA) ส่วนใหญ่เป็นยีสต์และจมูกข้าวสาลี ประเภทนี้มักถูกเรียกว่ากรดนิวคลีอิกของยีสต์
ในช่วงต้นทศวรรษ 1930 แนวคิดที่ว่าเซลล์พืชมีลักษณะเฉพาะด้วยกรดนิวคลีอิกประเภทยีสต์ค่อนข้างจะหยั่งรากอย่างแน่นหนา ในขณะที่กรดไทโมนิวคลีอิกมีลักษณะเฉพาะของนิวเคลียสของเซลล์สัตว์เท่านั้น กรดนิวคลีอิกสองประเภทคือ RNA และ DNA จากนั้นจึงเรียกกรดนิวคลีอิกจากพืชและสัตว์ตามลำดับ อย่างไรก็ตาม จากการศึกษาเบื้องต้นของ A.N. Belozersky พบว่าการแบ่งกรดนิวคลีอิกนั้นไม่ยุติธรรม ในปีพ.ศ. 2477 เบโลเซอร์สกีได้ค้นพบกรดไทโมนิวคลีอิกในเซลล์พืชเป็นครั้งแรก เขาแยกและระบุเบสไทมีน-ไพริมิดีนจากต้นกล้าถั่วจากต้นกล้าถั่ว จากนั้นเขาก็ค้นพบไทมีนในพืชชนิดอื่น (เมล็ดถั่วเหลือง ถั่ว) ในปี 1936 A. N. Belozersky และ I. I. Dubrovskaya แยก DNA ออกจากต้นกล้าเกาลัดม้า นอกจากนี้ ชุดของการศึกษาที่ดำเนินการในอังกฤษในทศวรรษที่ 1940 โดย D. Davidson และเพื่อนร่วมงานแสดงให้เห็นอย่างน่าเชื่อถือว่ากรดนิวคลีอิกจากพืช (RNA) มีอยู่ในเซลล์สัตว์หลายชนิด
การใช้ปฏิกิริยาไซโตเคมิคัลอย่างแพร่หลายสำหรับ DNA ที่พัฒนาโดย R. Felgen และ G. Rosenbeck (1924) และปฏิกิริยาของ J. Brachet (1944) สำหรับ RNA ทำให้สามารถแก้ไขปัญหาการแปลนิวเคลียสพิเศษได้อย่างรวดเร็วและชัดเจน กรดในเซลล์ ปรากฎว่า DNA มีความเข้มข้นในนิวเคลียส ในขณะที่ RNA นั้นมีความเข้มข้นอย่างเด่นชัดในไซโตพลาสซึม ต่อมาพบว่าอาร์เอ็นเอมีทั้งในไซโตพลาสซึมและในนิวเคลียสและนอกจากนี้ยังมีการระบุ DNA ของไซโตพลาสซึม
สำหรับคำถามเกี่ยวกับโครงสร้างหลักของกรดนิวคลีอิกในช่วงกลางทศวรรษ 1940 แนวคิดของ P. Levin ได้รับการจัดตั้งขึ้นอย่างมั่นคงในทางวิทยาศาสตร์ โดยที่กรดนิวคลีอิกทั้งหมดถูกสร้างขึ้นตามประเภทเดียวกันและประกอบด้วยเตตระนิวคลีโอไทด์ที่เหมือนกัน บล็อก แต่ละบล็อคเหล่านี้ ตามข้อมูลของ Lewin มีนิวคลีโอไทด์ที่แตกต่างกันสี่ชนิด ทฤษฎีเตตระนิวคลีโอไทด์ของโครงสร้างของกรดนิวคลีอิกส่วนใหญ่ทำให้ขาดความจำเพาะของพอลิเมอร์ชีวภาพเหล่านี้ ดังนั้นจึงไม่น่าแปลกใจที่ในเวลานั้นลักษณะเฉพาะของสิ่งมีชีวิตทั้งหมดเกี่ยวข้องกับโปรตีนเท่านั้นซึ่งธรรมชาติของโมโนเมอร์นั้นมีความหลากหลายมากกว่า (20 กรดอะมิโน)
ช่องว่างแรกในทฤษฎีโครงสร้างเตตระนิวคลีโอไทด์ของกรดนิวคลีอิกถูกสร้างขึ้นโดยข้อมูลการวิเคราะห์ของนักเคมีชาวอังกฤษ J. Gouland (1945 - 1947) เมื่อพิจารณาองค์ประกอบของกรดนิวคลีอิกโดยไนโตรเจนพื้นฐาน เขาไม่ได้รับอัตราส่วนสมมูลของเบสตามที่ควรจะเป็นตามทฤษฎีของเลวิน ในที่สุด ทฤษฎีเตตระนิวคลีโอไทด์ของโครงสร้างของกรดนิวคลีอิกก็พังทลายลงอันเป็นผลมาจากการวิจัยของอี. ชาร์กาฟฟ์และผู้ร่วมงานของเขา (พ.ศ. 2492 - 2494) Chargaff ใช้โครมาโตกราฟีแบบกระดาษเพื่อแยกเบสที่ปล่อยออกมาจาก DNA อันเป็นผลมาจากการไฮโดรไลซิสของกรดของมัน ฐานแต่ละฐานเหล่านี้ถูกกำหนดอย่างแม่นยำด้วยสเปกโตรโฟโตเมตริก Chargaff สังเกตเห็นความเบี่ยงเบนอย่างมีนัยสำคัญจากอัตราส่วนเท่ากันของเบสใน DNA ที่มีต้นกำเนิดต่างกัน และเป็นครั้งแรกที่กล่าวอย่างชัดเจนว่า DNA มีความจำเพาะของสายพันธุ์ที่เด่นชัด สิ่งนี้ทำให้ความเป็นเจ้าโลกของแนวคิดเรื่องความจำเพาะของโปรตีนในเซลล์ของสิ่งมีชีวิตสิ้นสุดลง การวิเคราะห์ DNA ของต้นกำเนิดที่แตกต่างกัน Chargaff ได้ค้นพบและกำหนดรูปแบบเฉพาะขององค์ประกอบ DNA ซึ่งเข้าสู่วิทยาศาสตร์ภายใต้ชื่อกฎของ Chargaff ตามกฎเหล่านี้ใน DNA ทั้งหมดโดยไม่คำนึงถึงแหล่งกำเนิดปริมาณของ adenine เท่ากับปริมาณของ thymine (A = T) ปริมาณของ guanine เท่ากับปริมาณของ cytosine (G = C) ปริมาณของ พิวรีนมีค่าเท่ากับปริมาณไพริมิดีน (G + A = C + T) ปริมาณเบสที่มีหมู่ 6 อะมิโนจะเท่ากับจำนวนเบสที่มีหมู่ 6-คีโต (A + C = G + T) ในเวลาเดียวกัน แม้ว่าจะมีการโต้ตอบเชิงปริมาณที่เข้มงวดเช่นนี้ แต่ DNA ของสปีชีส์ต่างๆ มีค่าของอัตราส่วน A+T:G+C ต่างกัน ใน DNA บางตัว ปริมาณของ guanine และ cytosine จะมีมากกว่าปริมาณของ adenine และ thymine (Chargaff เรียก DNA GC-type DNA เหล่านี้) DNA อื่นๆ มีอะดีนีนและไทมีนมากกว่ากัวนีนและไซโตซีน (DNA เหล่านี้เรียกว่า AT-type DNA) ข้อมูลที่ได้รับจาก Chargaff เกี่ยวกับองค์ประกอบของ DNA มีบทบาทพิเศษในด้านอณูชีววิทยา พวกมันเป็นพื้นฐานสำหรับการค้นพบโครงสร้างของ DNA ซึ่งสร้างในปี 1953 โดย J. Watson และ F. Crick
ย้อนกลับไปในปี 1938 W. Astbury และ F. Bell โดยใช้การวิเคราะห์การเลี้ยวเบนของรังสีเอกซ์ แสดงให้เห็นว่าระนาบฐานใน DNA ควรตั้งฉากกับแกนยาวของโมเลกุลและมีลักษณะคล้ายกับกองจานที่วางอยู่ด้านบน อื่น ๆ. ด้วยการปรับปรุงเทคนิคการวิเคราะห์การเลี้ยวเบนของรังสีเอกซ์ โดยปี พ.ศ. 2495 - พ.ศ. 2496 ข้อมูลที่สะสมทำให้สามารถตัดสินความยาวของพันธะแต่ละอันและมุมเอียงได้ สิ่งนี้ทำให้มีความเป็นไปได้มากที่สุดที่จะแสดงถึงธรรมชาติของการปฐมนิเทศของวงแหวนของสารตกค้างเพนโทสในกระดูกสันหลังน้ำตาลฟอสเฟตของโมเลกุลดีเอ็นเอ ในปีพ.ศ. 2495 เอส. ฟาร์เบิร์กได้เสนอแบบจำลองการเก็งกำไรของดีเอ็นเอสองแบบ ซึ่งเป็นตัวแทนของโมเลกุลสายเดี่ยวที่พับหรือบิดเบี้ยวด้วยตัวมันเอง แบบจำลองการเก็งกำไรไม่น้อยของโครงสร้างของ DNA ถูกเสนอในปี 1953 โดย L. Pauling (ผู้ชนะรางวัลโนเบล, 1954) และ R. Corey ในแบบจำลองนี้ DNA ที่บิดเป็นเกลียวสามเส้นก่อตัวเป็นเกลียวยาว ซึ่งแกนกลางของมันถูกแทนด้วยกลุ่มฟอสเฟต และฐานอยู่ด้านนอกของมัน ภายในปี 1953 เอ็ม. วิลกินส์และอาร์. แฟรงคลินได้รับรูปแบบการเลี้ยวเบนของดีเอ็นเอจากรังสีเอกซ์ที่ชัดเจนขึ้น การวิเคราะห์ของพวกเขาแสดงให้เห็นความล้มเหลวอย่างสมบูรณ์ของแบบจำลองของ Farberg, Pauling และ Corey จากการใช้ข้อมูลของ Chargaff เปรียบเทียบแบบจำลองโมเลกุลต่างๆ ของโมโนเมอร์แต่ละตัวและข้อมูลการเลี้ยวเบนของรังสีเอกซ์ J. Watson และ F. Crick ในปี 1953 ได้ข้อสรุปว่าโมเลกุลดีเอ็นเอจะต้องเป็นเกลียวคู่ กฎของ Chargaff จำกัดจำนวนของการรวมฐานที่เป็นไปได้อย่างมากในแบบจำลองดีเอ็นเอที่เสนอ พวกเขาแนะนำวัตสันและคริกว่าต้องมีการจับคู่เบสเฉพาะในโมเลกุลดีเอ็นเอ - อะดีนีนกับไทมีนและกวานีนกับไซโตซีน กล่าวอีกนัยหนึ่ง อะดีนีนในดีเอ็นเอสายหนึ่งจะสัมพันธ์กับไทมีนในสายอื่นอย่างเคร่งครัด และกวานีนในสายหนึ่งจำเป็นต้องสอดคล้องกับไซโตซีนในอีกสายหนึ่ง ดังนั้น วัตสันและคริกจึงได้กำหนดหลักการของโครงสร้างเสริมของ DNA ที่มีความสำคัญเป็นพิเศษเป็นครั้งแรก โดยที่ DNA สายหนึ่งไปประกอบกับอีกสายหนึ่ง นั่นคือ ลำดับเบสของสายหนึ่งเป็นตัวกำหนดลำดับของเบสในอีกสายหนึ่งโดยเฉพาะ (ส่วนเสริม) ) สาระ เห็นได้ชัดว่าในโครงสร้างของ DNA มีศักยภาพในการสืบพันธุ์ที่แน่นอนอยู่แล้ว แบบจำลองโครงสร้างดีเอ็นเอนี้เป็นที่ยอมรับกันโดยทั่วไปในปัจจุบัน Crick, Watson และ Wilkins ได้รับรางวัลโนเบลในปี 1962 จากการถอดรหัสโครงสร้างของ DNA
ควรสังเกตว่าแนวคิดของกลไกในการทำซ้ำของโมเลกุลขนาดใหญ่และการส่งข้อมูลทางพันธุกรรมที่เกิดขึ้นในประเทศของเรา ในปี 1927 N. K. Koltsov เสนอแนะว่าในระหว่างการสืบพันธุ์ของเซลล์ การสืบพันธุ์ของโมเลกุลเกิดขึ้นจากการสืบพันธุ์แบบอัตโนมัติของโมเลกุลต้นกำเนิดที่มีอยู่ จริงอยู่ในเวลานั้น Koltsov มอบคุณสมบัตินี้ไม่ใช่ด้วยโมเลกุล DNA แต่ด้วยโมเลกุลที่มีลักษณะโปรตีนซึ่งไม่ทราบถึงความสำคัญเชิงหน้าที่ อย่างไรก็ตามแนวคิดของการสืบพันธุ์แบบอัตโนมัติของโมเลกุลขนาดใหญ่และกลไกการถ่ายทอดคุณสมบัติทางพันธุกรรมกลายเป็นคำทำนาย: มันกลายเป็นแนวความคิดของชีววิทยาโมเลกุลสมัยใหม่
ดำเนินการในห้องปฏิบัติการของ A. N. Belozersky โดย A. S. Spirin, G. N. Zaitseva, B. F. Vanyushin, S. O. Uryson, A. S. Antonov และสิ่งมีชีวิตที่หลากหลายอื่น ๆ ได้ยืนยันรูปแบบที่ Chargaff ค้นพบอย่างเต็มที่และปฏิบัติตามแบบจำลองโมเลกุลของโครงสร้างของ DNA ที่เสนอโดย Watson อย่างสมบูรณ์ และคริก การศึกษาเหล่านี้แสดงให้เห็นว่า DNA ของแบคทีเรีย เชื้อรา สาหร่าย แอกติโนมัยซีต พืชชั้นสูง สัตว์ไม่มีกระดูกสันหลัง และสัตว์มีกระดูกสันหลังที่แตกต่างกันมีองค์ประกอบเฉพาะ ความแตกต่างในองค์ประกอบ (เนื้อหาของคู่ AT-base) นั้นเด่นชัดเป็นพิเศษในจุลินทรีย์ กลายเป็นคุณสมบัติการจัดอนุกรมวิธานที่สำคัญ ในพืชและสัตว์ชั้นสูง การแปรผันของสปีชีส์ในองค์ประกอบของดีเอ็นเอนั้นเด่นชัดน้อยกว่ามาก แต่นี่ไม่ได้หมายความว่า DNA ของพวกเขามีความเฉพาะเจาะจงน้อยกว่า นอกจากองค์ประกอบของเบสแล้ว ความจำเพาะส่วนใหญ่ถูกกำหนดโดยลำดับของเบสในสายดีเอ็นเอ
นอกจากเบสปกติแล้ว ยังพบเบสไนโตรเจนเพิ่มเติมใน DNA และ RNA ดังนั้น G. White (1950) จึงพบ 5-methylcytosine ใน DNA ของพืชและสัตว์ และ D. Dunn และ J. Smith (1958) พบ methylated adenine ใน DNA บางตัว เป็นเวลานาน methylcytosine ถือเป็นจุดเด่นของสารพันธุกรรมของสิ่งมีชีวิตชั้นสูง ในปี 1968 A. N. Belozersky, B. F. Vanyushin และ N. A. Kokurina พบว่าสามารถพบเชื้อนี้ใน DNA ของแบคทีเรียได้เช่นกัน
ในปีพ.ศ. 2507 M. Gold และ J. Hurwitz ได้ค้นพบเอ็นไซม์ชนิดใหม่ที่ทำการดัดแปลงดีเอ็นเอตามธรรมชาติ - เมทิลเลชันของมัน หลังจากการค้นพบนี้ เป็นที่แน่ชัดว่าเบสรอง (ที่มีอยู่ในปริมาณเล็กน้อย) เกิดขึ้นแล้วบนสายโซ่พอลินิวคลีโอไทด์ของ DNA ที่เสร็จสิ้นแล้วอันเป็นผลมาจากเมทิเลชันจำเพาะของไซโตซีนและสารตกค้างของอะดีนีนในลำดับพิเศษ โดยเฉพาะอย่างยิ่งตาม B. F. Vanyushin, Ya. I. Buryanov และ A. N. Belozersky (1969), adenine methylation ใน Escherichia coli DNA สามารถเกิดขึ้นได้ในการยุติ codons ตามที่ A.N. Belozersky และเพื่อนร่วมงาน (1968 - 1970) เช่นเดียวกับ M. Meselson (USA) และ V. Arber (สวิตเซอร์แลนด์) (1965 - 1969) methylation ให้คุณสมบัติเฉพาะตัวของโมเลกุล DNA และเมื่อรวมกับการกระทำของ นิวคลีเอสจำเพาะเป็นส่วนหนึ่งของกลไกที่ซับซ้อนที่ควบคุมการสังเคราะห์ DNA ในเซลล์ กล่าวอีกนัยหนึ่ง ธรรมชาติของการเกิดเมทิลเลชันของ DNA โดยเฉพาะกำหนดคำถามว่าจะสามารถเพิ่มจำนวนในเซลล์ที่กำหนดได้หรือไม่
เกือบจะในเวลาเดียวกัน การแยกและการศึกษาอย่างเข้มข้นของ DNA methylases และ endonucleases ข้อจำกัดก็เริ่มขึ้น ในปี 2512 - 2518 ลำดับนิวคลีโอไทด์ที่รู้จักใน DNA โดยเอ็นไซม์เหล่านี้บางตัวได้ถูกสร้างขึ้น (X. Boyer, X. Smith, S. Lynn, K. Murray) เมื่อดีเอ็นเอต่างๆ ถูกไฮโดรไลซ์ด้วยเอ็นไซม์จำกัด ชิ้นส่วนที่ค่อนข้างใหญ่ที่มีปลาย "เหนียว" เหมือนกันจะถูกตัดออก ทำให้ไม่เพียงแต่วิเคราะห์โครงสร้างของยีน เช่นเดียวกับที่ทำในไวรัสขนาดเล็ก (D. Nathans, S. Adler, 1973 - 1975) แต่ยังสร้างจีโนมต่างๆ ได้อีกด้วย ด้วยการค้นพบเอ็นไซม์จำกัดจำเพาะเหล่านี้ พันธุวิศวกรรมได้กลายเป็นความจริงที่จับต้องได้ ที่ฝังอยู่ในยีน DNA พลาสมิดขนาดเล็กที่มีต้นกำเนิดต่างๆ ได้ถูกนำเข้ามาอย่างง่ายดายในเซลล์ต่างๆ ดังนั้นจึงได้พลาสมิดที่ออกฤทธิ์ทางชีวภาพชนิดใหม่ ทำให้ดื้อต่อยาปฏิชีวนะบางชนิด (S. Cohen, 1973), ยีนไรโบโซมของกบและแมลงหวี่ถูกนำเข้าสู่ Escherichia coli plasmids (J. Morrow, 1974; X. Boyer, D . ฮอกเนส, อาร์. เดวิส , 2517 - 2518). ดังนั้น วิธีการที่แท้จริงจึงเปิดกว้างสำหรับการได้รับสิ่งมีชีวิตใหม่โดยพื้นฐานโดยการแนะนำและบูรณาการยีนต่างๆ ลงในกลุ่มยีนของพวกมัน การค้นพบนี้สามารถนำไปสู่ประโยชน์ของมนุษยชาติทั้งหมด
ในปี 1952 G. White และ S. Cohen ค้นพบว่า DNA ของ T-even phages มีเบสที่ผิดปกติคือ 5-hydroxymethylcytosine ต่อมาจากผลงานของ E. Volkin และ R. Sinsheimer (1954) และ Cohen (1956) เป็นที่ทราบกันดีอยู่แล้วว่าสารตกค้างของ hydroxymethylcytosine สามารถถูกกลูโคซิไดซ์ทั้งหมดหรือบางส่วนได้ ซึ่งเป็นผลมาจากการที่โมเลกุล DNA ของ phage ได้รับการปกป้องจากปฏิกิริยาไฮโดรไลติก ของนิวคลีเอส
ในช่วงต้นทศวรรษ 1950 จากผลงานของ D. Dunn และ J. Smith (อังกฤษ), S. Zamenhof (USA) และ A. Wacker (เยอรมนี) เป็นที่ทราบกันดีว่าสารอะนาล็อกพื้นฐานเทียมจำนวนมากสามารถรวมเข้ากับ DNA ได้ ซึ่งบางครั้งก็มาแทนที่ ไทมีนสูงถึง 50% ตามกฎแล้ว การแทนที่เหล่านี้จะนำไปสู่ข้อผิดพลาดในการจำลองแบบ DNA การถอดความและการแปล และการปรากฏตัวของการกลายพันธุ์ ดังนั้น J. Marmur (1962) พบว่า DNA ของฟาจบางตัวมี oxymethyluracil แทนที่จะเป็นไทมีน ในปี 1963 I. Takahashi และ J. Marmur ค้นพบว่า DNA ของฟาจตัวใดตัวหนึ่งมียูราซิลแทนที่จะเป็นไทมีน ดังนั้นหลักการอื่นตามที่กรดนิวคลีอิกแยกจากกันก่อนหน้านี้จึงพังทลายลง ตั้งแต่เวลาของการทำงานของ P. Levin เป็นที่เชื่อกันว่าไทมีนเป็นจุดเด่นของ DNA และ uracil เป็นจุดเด่นของ RNA เป็นที่ชัดเจนว่าสัญลักษณ์นี้ไม่น่าเชื่อถือเสมอไป และความแตกต่างพื้นฐานในลักษณะทางเคมีของกรดนิวคลีอิกสองประเภทดังที่ปรากฏในปัจจุบันเป็นเพียงลักษณะขององค์ประกอบคาร์โบไฮเดรตเท่านั้น
ในการศึกษาฟาจ ได้เปิดเผยลักษณะพิเศษหลายอย่างของการจัดระเบียบของกรดนิวคลีอิก ตั้งแต่ปี 1953 เป็นที่เชื่อกันว่า DNA ทั้งหมดเป็นโมเลกุลเชิงเส้นตรงที่มีสายคู่ ในขณะที่ RNA นั้นมีสายเพียงเส้นเดียว ตำแหน่งนี้สั่นสะเทือนอย่างมีนัยสำคัญในปี 1961 เมื่อ R. Sinsheimer ค้นพบว่า DNA ของ phage φ X 174 นั้นแสดงด้วยโมเลกุลวงกลมเส้นเดียว อย่างไรก็ตาม ภายหลังปรากฎว่าในรูปแบบนี้ DNA นี้มีอยู่ในอนุภาคฟาจจากพืชเท่านั้น และรูปแบบการจำลองแบบของ DNA ของฟาจนี้ก็เป็นแบบเกลียวคู่เช่นกัน นอกจากนี้ กลับกลายเป็นว่าค่อนข้างไม่คาดคิดว่า RNA ของไวรัสบางตัวสามารถเป็นแบบสองสายได้ การจัดระเบียบโมเลกุลของอาร์เอ็นเอรูปแบบใหม่นี้ถูกค้นพบในปี 2505 โดย P. Gomatos, I. Tamm และนักวิจัยคนอื่น ๆ ในไวรัสสัตว์บางชนิดและในไวรัสเนื้องอกบาดแผลในพืช เมื่อเร็ว ๆ นี้ V. I. Agol และ A. A. Bogdanov (1970) ได้กำหนดว่านอกเหนือจากโมเลกุล RNA เชิงเส้นแล้วยังมีโมเลกุลแบบปิดหรือแบบวัฏจักรอีกด้วย พวกเขาตรวจพบ RNA แบบเกลียวคู่แบบวัฏจักรโดยเฉพาะอย่างยิ่งในไวรัสไข้สมองอักเสบ ขอบคุณผลงานของ X. Deveaux, L. Tinoko, T. I. Tikhonenko, E. I. Budovsky และคนอื่น ๆ (1960 - 1974) คุณสมบัติหลักขององค์กร (การวาง) ของสารพันธุกรรมในแบคทีเรียกลายเป็นที่รู้จัก
ในช่วงปลายทศวรรษ 1950 นักวิทยาศาสตร์ชาวอเมริกัน P. Doty พบว่าการให้ความร้อนทำให้เกิด DNA denaturation ซึ่งมาพร้อมกับการทำลายพันธะไฮโดรเจนระหว่างคู่เบสกับการแยกสายโซ่เสริม กระบวนการนี้มีลักษณะของการเปลี่ยนเฟส "ขดลวดเกลียว" และคล้ายกับการละลายของผลึก ดังนั้น Doty จึงเรียกกระบวนการเปลี่ยนสภาพด้วยความร้อนของการหลอม DNA DNA ด้วยการเย็นตัวช้าจะเกิดการเปลี่ยนสีของโมเลกุลนั่นคือการรวมตัวของส่วนเสริม
หลักการของการเปลี่ยนสภาพในปี 2503 ถูกใช้โดย J. Marmur และ K. Schildkraut เพื่อกำหนดระดับของ "ความสามารถในการผสมพันธุ์" ของ DNA ของจุลินทรีย์ต่างๆ ต่อจากนั้น E. Bolton และ B. McCarthy ได้ปรับปรุงเทคนิคนี้โดยเสนอวิธีการที่เรียกว่าคอลัมน์ DNA-agar วิธีนี้กลายเป็นสิ่งที่ขาดไม่ได้ในการศึกษาระดับความคล้ายคลึงกันของลำดับนิวคลีโอไทด์ของ DNA ต่างๆ และชี้แจงความสัมพันธ์ทางพันธุกรรมของสิ่งมีชีวิตต่างๆ การสลายตัวของดีเอ็นเอที่ค้นพบโดย Doty ร่วมกับโครมาโตกราฟีบนอัลบูมินที่มีเมทิลที่บรรยายโดย J. Mandel และ A. Hershey * (1960) และการหมุนเหวี่ยงเกรเดียนต์ของความหนาแน่น (วิธีการนี้พัฒนาขึ้นในปี 2500 โดย M. Meselson, F. Stahl และ D. Winograd) ใช้กันอย่างแพร่หลายสำหรับการแยก การแยก และการวิเคราะห์สาย DNA เสริมแต่ละเส้น ตัวอย่างเช่น W. Shibalsky (USA) โดยใช้เทคนิคเหล่านี้เพื่อแยก DNA ของ lambda phage ออกแสดงในปี 1967 - 1969 ว่าสายโซ่ฟาจทั้งสองมีการเคลื่อนไหวทางพันธุกรรม และไม่ใช่อย่างใดอย่างหนึ่ง เนื่องจากสิ่งนี้ถือเป็น (S. Spiegelman, 1961). ควรสังเกตว่าเป็นครั้งแรกที่ความคิดเกี่ยวกับความสำคัญทางพันธุกรรมของสาย DNA ทั้งสองของแลมบ์ดาฟาจถูกแสดงออกในสหภาพโซเวียตโดย SE Bresler (1961)
* (สำหรับงานของพวกเขาเกี่ยวกับพันธุกรรมของแบคทีเรียและไวรัส A. Hershey ร่วมกับ M. Delbrück และ S. Luria ได้รับรางวัลโนเบลในปี 1969)
เพื่อให้เข้าใจถึงการจัดระเบียบและกิจกรรมการทำงานของจีโนม การกำหนดลำดับดีเอ็นเอนิวคลีโอไทด์มีความสำคัญอย่างยิ่ง การค้นหาวิธีการของความมุ่งมั่นดังกล่าวดำเนินการในห้องปฏิบัติการหลายแห่งทั่วโลก นับตั้งแต่ช่วงปลายทศวรรษ 1950 M. Beer และผู้ทำงานร่วมกันได้พยายามสร้างลำดับดีเอ็นเอโดยใช้กล้องจุลทรรศน์อิเล็กตรอนในสหรัฐอเมริกา แต่จนถึงขณะนี้ไม่ประสบความสำเร็จ ในช่วงต้นทศวรรษ 1950 จากงานแรกของ Sinsheimer, Chargaff และนักวิจัยคนอื่นๆ เกี่ยวกับการย่อยสลายของเอนไซม์ของ DNA เป็นที่ทราบกันดีว่ามีการกระจายนิวคลีโอไทด์ที่แตกต่างกันในโมเลกุล DNA แม้ว่าจะไม่ได้สุ่มแต่ก็ไม่สม่ำเสมอ ตามที่นักเคมีชาวอังกฤษ C. Barton (1961) ระบุว่า pyrimidines (มากกว่า 70%) มีความเข้มข้นส่วนใหญ่อยู่ในรูปแบบของบล็อกที่เกี่ยวข้อง A. L. Mazin และ B.F. Vanyushin (1968 - 1969) พิสูจน์ว่า DNA ที่แตกต่างกันมีระดับการเกาะติดกันของ pyrimidine ต่างกัน และใน DNA ของสิ่งมีชีวิตสัตว์นั้นจะเพิ่มขึ้นอย่างเห็นได้ชัดเมื่อเคลื่อนจากล่างขึ้นสู่สูง ดังนั้นวิวัฒนาการของสิ่งมีชีวิตจึงสะท้อนให้เห็นในโครงสร้างของจีโนมของพวกมันด้วย ด้วยเหตุนี้ สำหรับการทำความเข้าใจกระบวนการวิวัฒนาการโดยรวม การศึกษาเปรียบเทียบโครงสร้างของกรดนิวคลีอิกจึงมีความสำคัญเป็นพิเศษ การวิเคราะห์โครงสร้างของพอลิเมอร์ที่มีความสำคัญทางชีววิทยา และประการแรก DNA มีความสำคัญอย่างยิ่งในการแก้ปัญหาเฉพาะจำนวนมากของสายวิวัฒนาการและอนุกรมวิธาน
เป็นที่น่าสนใจที่จะสังเกตว่านักสรีรวิทยาชาวอังกฤษ อี. แลงเคสเตอร์ ผู้ศึกษาฮีโมโกลบินของหอย คาดการณ์แนวคิดของอณูชีววิทยาเมื่อ 100 ปีที่แล้ว เขียนว่า: “ความแตกต่างทางเคมีระหว่างสปีชีส์และสกุลของสัตว์และพืชต่างกันมีความสำคัญต่อการอธิบายให้กระจ่าง ประวัติต้นกำเนิดของมันเป็นรูปแบบของพวกมันหากเราสามารถระบุความแตกต่างในการจัดระเบียบระดับโมเลกุลและการทำงานของสิ่งมีชีวิตได้อย่างชัดเจนเราจะสามารถเข้าใจกำเนิดและวิวัฒนาการของสิ่งมีชีวิตต่าง ๆ ได้ดีกว่าการสังเกตทางสัณฐานวิทยา " * . ความสำคัญของการศึกษาทางชีวเคมีสำหรับอนุกรมวิธานยังเน้นย้ำโดย V. L. Komarov ผู้เขียนว่า "พื้นฐานของลักษณะทางสัณฐานวิทยาล้วนๆ ทั้งหมด บนพื้นฐานของการที่เราจำแนกและสร้างสปีชีส์นั้นมีความแตกต่างทางชีวเคมีอย่างแม่นยำ" **
* (อี.อาร์.แลงเคสเตอร์ Uber das Vorcommen von Hemoglobin ใน den Muskeln der Mollusken und die Verbreitung desselben in den lebendigen Organismen.- "Pfluger" s Archiv fur die gesammte Physiol., 1871, Bd 4, 319.)
** (วี.แอล.โคมารอฟ. ผลงานที่เลือก เล่ม 1 M.-L. สำนักพิมพ์ Academy of Sciences of the USSR, 1945, p. 331)
A. V. Blagoveshchenskii และ S. L. Ivanov ย้อนกลับไปในปี ค.ศ. 1920 ได้ใช้ขั้นตอนแรกในประเทศของเราเพื่อชี้แจงคำถามบางประการเกี่ยวกับวิวัฒนาการและการจัดระบบของสิ่งมีชีวิตบนพื้นฐานของการวิเคราะห์เปรียบเทียบองค์ประกอบทางชีวเคมีของพวกมัน (ดูบทที่ 2) การวิเคราะห์เปรียบเทียบโครงสร้างของโปรตีนและกรดนิวคลีอิกได้กลายเป็นเครื่องมือที่จับต้องได้มากขึ้นสำหรับนักอนุกรมวิธาน (ดูบทที่ 21) วิธีการของอณูชีววิทยานี้ทำให้ไม่เพียงแต่ชี้แจงตำแหน่งของแต่ละสปีชีส์ในระบบเท่านั้น แต่ยังทำให้จำเป็นต้องพิจารณาถึงหลักการของการจำแนกสิ่งมีชีวิตใหม่ ๆ และบางครั้งก็แก้ไขทั้งระบบโดยรวม เช่นที่เกิดขึ้นกับระบบจุลินทรีย์ ไม่ต้องสงสัยเลย ในอนาคต การวิเคราะห์โครงสร้างของจีโนมจะเป็นศูนย์กลางในการจัดระบบเคมีของสิ่งมีชีวิต
ความสำคัญอย่างยิ่งต่อการพัฒนาของอณูชีววิทยาคือการถอดรหัสกลไกของการจำลองแบบดีเอ็นเอและการถอดรหัส (ดูบทที่ 24)
การสังเคราะห์โปรตีน
การเปลี่ยนแปลงที่สำคัญในการแก้ปัญหาการสังเคราะห์โปรตีนเกี่ยวข้องกับความก้าวหน้าในการศึกษากรดนิวคลีอิก ในปี 1941 T. Kasperson (สวีเดน) และในปี 1942 J. Brachet (เบลเยียม) ให้ความสนใจกับความจริงที่ว่าเนื้อเยื่อที่มีการสังเคราะห์โปรตีนเชิงรุกมีปริมาณ RNA เพิ่มขึ้น พวกเขาสรุปว่ากรดไรโบนิวคลีอิกมีบทบาทสำคัญในการสังเคราะห์โปรตีน ในปี 1953 E. Gale และ D. Fox ดูเหมือนจะได้รับหลักฐานโดยตรงถึงการมีส่วนร่วมโดยตรงของ RNA ในการสังเคราะห์โปรตีน: จากข้อมูลของพวกเขา ribonuclease ยับยั้งการรวมตัวของกรดอะมิโนในเซลล์แบคทีเรีย lysates อย่างมีนัยสำคัญ ข้อมูลที่คล้ายกันได้มาจาก V. Olfri, M. Delhi และ A. Mirsky (1953) เกี่ยวกับตับที่เป็นเนื้อเดียวกัน ต่อมา E. Gale ปฏิเสธความคิดที่ถูกต้องของเขาเกี่ยวกับบทบาทนำของ RNA ในการสังเคราะห์โปรตีน โดยเชื่ออย่างผิด ๆ ว่าการกระตุ้นการสังเคราะห์โปรตีนในระบบที่ปราศจากเซลล์นั้นเกิดขึ้นภายใต้อิทธิพลของสารอื่นที่ไม่ทราบธรรมชาติ ในปี 1954 P. Zamechnik, D. Littlefield, R. B. Khesin-Lurie และคนอื่น ๆ พบว่าการรวมตัวของกรดอะมิโนที่แอคทีฟมากที่สุดเกิดขึ้นในเศษส่วนของอนุภาคย่อยเซลล์ - ไมโครโซมที่อุดมด้วย RNA P. Zamechnik และ E. Keller (1953 - 1954) พบว่าการรวมตัวของกรดอะมิโนได้รับการปรับปรุงอย่างเห็นได้ชัดเมื่อมี supernatant ภายใต้สภาวะของการสร้าง ATP ขึ้นใหม่ P. Sikevitz (1952) และ M. Hoagland (1956) แยกส่วนของโปรตีน (เศษส่วน pH 5) ออกจาก supernatant ซึ่งมีหน้าที่ในการกระตุ้นการรวมกรดอะมิโนในไมโครโซมอย่างคมชัด นอกจากโปรตีนแล้ว ยังพบ RNA ที่มีน้ำหนักโมเลกุลต่ำซึ่งปัจจุบันเรียกว่า transfer RNAs (tRNAs) ในระดับพิเศษใน supernatant ในปี 1958 Hoagland และ Zamechnik รวมถึง P. Berg, R. Sweet และ F. Allen และนักวิจัยอีกหลายคนพบว่ากรดอะมิโนแต่ละชนิดต้องการเอนไซม์พิเศษ ATP และ tRNA เฉพาะของตัวเองเพื่อกระตุ้น เป็นที่ชัดเจนว่า tRNA ทำหน้าที่เฉพาะของอะแดปเตอร์ เช่น อุปกรณ์ที่หาตำแหน่งบนนิวคลีอิกเมทริกซ์ (mRNA) สำหรับกรดอะมิโนที่สอดคล้องกันในโมเลกุลโปรตีนที่เกิดขึ้นใหม่ การศึกษาเหล่านี้ยืนยันสมมติฐานอะแดปเตอร์ของ F. Crick (1957) อย่างเต็มที่ซึ่งมีให้สำหรับการมีอยู่ของตัวปรับต่อพอลินิวคลีโอไทด์ในเซลล์ ซึ่งจำเป็นสำหรับการจัดเรียงที่ถูกต้องของกรดอะมิโนตกค้างของโปรตีนสังเคราะห์บนเมทริกซ์นิวคลีอิก ต่อมา นักวิทยาศาสตร์ชาวฝรั่งเศส F. Chapville (1962) ในห้องทดลองของ F. Lipman (รางวัลโนเบล, 1953) ในสหรัฐอเมริกาอย่างชาญฉลาดและชัดเจนแสดงให้เห็นอย่างชัดเจนว่าตำแหน่งของกรดอะมิโนในโมเลกุลโปรตีนสังเคราะห์ถูกกำหนดโดยสมบูรณ์ tRNA เฉพาะที่ติดอยู่ สมมติฐานอะแดปเตอร์ของ Crick ได้รับการพัฒนาโดย Hoagland และ Zamechnik
ในปี 1958 ระยะหลักต่อไปนี้ของการสังเคราะห์โปรตีนกลายเป็นที่รู้จัก: 1) การกระตุ้นกรดอะมิโนโดยเอนไซม์เฉพาะจาก "เศษส่วน pH 5" ต่อหน้า ATP ด้วยการก่อตัวของอะมิโนอะซิลอะดีนิเลต; 2) การแนบกรดอะมิโนที่ถูกกระตุ้นเข้ากับ tRNA เฉพาะด้วยการปล่อยอะดีโนซีนโมโนฟอสเฟต (AMP) 3) การจับกันของ aminoacyl-tRNA (tRNA ที่เต็มไปด้วยกรดอะมิโน) กับไมโครโซมและการรวมตัวของกรดอะมิโนเข้ากับโปรตีนด้วยการปล่อย tRNA Hoagland (1958) ตั้งข้อสังเกตว่า guanosine triphosphate (GTP) เป็นสิ่งจำเป็นในขั้นตอนสุดท้ายของการสังเคราะห์โปรตีน
ถ่ายโอน RNA และการสังเคราะห์ยีน
หลังจากการค้นพบ tRNAs การค้นหาการแยกส่วนและการกำหนดลำดับนิวคลีโอไทด์เริ่มต้นขึ้น นักชีวเคมีชาวอเมริกัน R. Holly ประสบความสำเร็จอย่างสูงสุด ในปีพ.ศ. 2508 เขาได้สร้างโครงสร้างของอะลานีน tRNA จากยีสต์ การใช้ไรโบนิวคลีเอส (guanyl RNase และ RNase ของตับอ่อน) ฮอลลี่แบ่งโมเลกุลกรดนิวคลีอิกออกเป็นหลายส่วน กำหนดลำดับนิวคลีโอไทด์ในแต่ละส่วนแยกกัน จากนั้นจึงสร้างลำดับของโมเลกุลอะลานีน tRNA ทั้งหมดขึ้นใหม่ วิธีการวิเคราะห์ลำดับนิวคลีโอไทด์นี้เรียกว่าวิธีบล็อก ข้อดีของ Holly ส่วนใหญ่ประกอบด้วยความจริงที่ว่าเขาเรียนรู้ที่จะแบ่งโมเลกุล RNA ไม่เพียง แต่เป็นชิ้นเล็ก ๆ อย่างที่หลายคนทำก่อนหน้าเขา แต่ยังเป็นชิ้นใหญ่ (ไตรมาสและครึ่งหนึ่ง) สิ่งนี้ทำให้เขามีโอกาสประกอบชิ้นส่วนเล็กๆ แต่ละชิ้นเข้าด้วยกันอย่างถูกต้อง และด้วยเหตุนี้จึงสร้างลำดับนิวคลีโอไทด์ที่สมบูรณ์ของโมเลกุล tRNA ทั้งหมด (รางวัลโนเบล, 1968)
ห้องปฏิบัติการหลายแห่งทั่วโลกใช้เทคนิคนี้ในทันที ในอีกสองปีข้างหน้า โครงสร้างหลักของ tRNA หลายตัวถูกถอดรหัสในสหภาพโซเวียตและในต่างประเทศ A. A. Baev (1967) และเพื่อนร่วมงานได้สร้างลำดับนิวคลีโอไทด์ในยีสต์ valine tRNA เป็นครั้งแรก จนถึงปัจจุบัน มีการศึกษา tRNA ที่แตกต่างกันมากกว่าหนึ่งโหล บันทึกที่แปลกประหลาดในการกำหนดลำดับนิวคลีโอไทด์ถูกตั้งค่าในเคมบริดจ์โดย F. Senger และ G. Brownlee นักวิจัยเหล่านี้ได้พัฒนาวิธีการแยกโอลิโกนิวคลีโอไทด์และจัดลำดับ RNA 5 S (ไรโบโซม) ที่เรียกกันว่า 5 S (ไรโบโซม) ออกจากเซลล์ E. coli ได้อย่างสง่างามอย่างน่าประหลาดใจ RNA นี้ประกอบด้วย 120 นิวคลีโอไทด์เรซิดิว และไม่เหมือนกับ tRNA ที่ไม่มีเบสรองเพิ่มเติม ซึ่งอำนวยความสะดวกในการวิเคราะห์ลำดับนิวคลีโอไทด์อย่างมาก โดยทำหน้าที่เป็นจุดสังเกตที่ไม่ซ้ำกันสำหรับชิ้นส่วนแต่ละชิ้นของโมเลกุล ในปัจจุบัน ต้องขอบคุณการใช้วิธีการของแซงเกอร์และบราวน์ลี การศึกษาลำดับของไรโบโซมอาร์เอ็นเอแบบยาวและอาร์เอ็นเอของไวรัสบางชนิดกำลังก้าวหน้าอย่างประสบความสำเร็จในห้องปฏิบัติการของเจ. เอเบล (ฝรั่งเศส) และนักวิจัยคนอื่นๆ
A. A. Baev และคณะ (1967) พบว่า valine tRNA ที่ผ่าครึ่งช่วยฟื้นฟูโครงสร้างโมเลกุลขนาดใหญ่ในสารละลาย และถึงแม้จะมีข้อบกพร่องในโครงสร้างหลัก แต่ก็มีกิจกรรมการทำงานของโมเลกุลดั้งเดิม (ดั้งเดิม) วิธีการนี้ - การสร้างโมเลกุลขนาดใหญ่ขึ้นใหม่หลังจากการกำจัดเศษบางส่วน - กลับกลายเป็นว่ามีแนวโน้มมาก ปัจจุบันมีการใช้กันอย่างแพร่หลายเพื่ออธิบายบทบาทหน้าที่ของแต่ละส่วนของ tRNA บางตัว
ในช่วงไม่กี่ปีที่ผ่านมา ประสบความสำเร็จอย่างมากในการเตรียมผลึกของ tRNA แต่ละตัว tRNA จำนวนมากได้รับการตกผลึกแล้วในห้องปฏิบัติการหลายแห่งในสหรัฐอเมริกาและอังกฤษ ทำให้สามารถศึกษาโครงสร้างของ tRNA โดยใช้การวิเคราะห์การเลี้ยวเบนของรังสีเอกซ์ ในปี 1970 R. Bock ได้นำเสนอรูปแบบ X-ray แรกและแบบจำลองสามมิติของ tRNA หลายตัวที่เขาสร้างขึ้นที่มหาวิทยาลัยวิสคอนซิน โมเดลเหล่านี้ช่วยกำหนดโลคัลไลเซชันของไซต์ที่ทำงานตามหน้าที่แต่ละแห่งใน tRNA และเข้าใจหลักการพื้นฐานของการทำงานของโมเลกุลเหล่านี้
การถอดรหัสธรรมชาติของรหัสพันธุกรรม (ดูบทที่ 24) ซึ่งถือได้ว่าเป็นความสำเร็จระดับแนวหน้าของวิทยาศาสตร์ธรรมชาติในศตวรรษที่ 20 โดยปราศจากการพูดเกินจริง มีความสำคัญอย่างยิ่งต่อการเปิดเผยกลไกการสังเคราะห์โปรตีนและการแก้ปัญหา ของความจำเพาะของกระบวนการนี้
การค้นพบโครงสร้างหลักของ tRNA ของ R. Holly ทำให้เกิดแรงผลักดันให้งานของ G. Korana * (USA) ในการสังเคราะห์โอลิโกนิวคลีโอไทด์และชี้นำพวกมันไปสู่การสังเคราะห์โครงสร้างทางชีววิทยาจำเพาะ - โมเลกุล DNA ที่เข้ารหัสอะลานีน tRNA ขั้นตอนแรกในการสังเคราะห์ทางเคมีของโอลิโกนิวคลีโอไทด์สั้นที่ทำโดยอัลกุรอานเมื่อเกือบ 15 ปีที่แล้วสิ้นสุดลงในปี 2513 ด้วยการสังเคราะห์ยีนครั้งแรก อัลกุรอานและผู้ทำงานร่วมกันของเขาได้สังเคราะห์ชิ้นส่วนสั้น ๆ ของนิวคลีโอไทด์ 8-12 ชิ้นจากนิวคลีโอไทด์แต่ละตัว ชิ้นส่วนเหล่านี้ที่มีลำดับนิวคลีโอไทด์ที่กำหนดให้ก่อตัวเป็นชิ้นส่วนประกอบแบบเกลียวคู่ที่เกิดขึ้นเองโดยมีการทับซ้อนกันของนิวคลีโอไทด์ 4-5 ตัว จากนั้นชิ้นงานสำเร็จรูปเหล่านี้ถูกต่อเข้าด้วยกันในลำดับที่ถูกต้องโดยใช้เอนไซม์ DNA ligase ดังนั้น ตรงกันข้ามกับการจำลองแบบของโมเลกุลดีเอ็นเอ ตาม A. Kornberg ** (ดูบทที่ 24) คัมภีร์กุรอ่านสามารถสร้างโมเลกุลดีเอ็นเอสายคู่ตามธรรมชาติขึ้นใหม่ตามโปรแกรมที่วางแผนไว้ล่วงหน้าตาม ลำดับ tRNA ที่ฮอลลี่อธิบาย ในทำนองเดียวกัน ขณะนี้งานกำลังดำเนินการเกี่ยวกับการสังเคราะห์ยีนอื่นๆ (M. N. Kolosov, Z. A. Shabarova, D. G. Knorre, 1970 - 1975)
* (สำหรับการศึกษารหัสพันธุกรรม G. Koran และ M. Nirenberg ได้รับรางวัลโนเบลในปี 1968)
** (สำหรับการค้นพบโพลีเมอเรสและการสังเคราะห์ DNA A. Kornberg และสำหรับการสังเคราะห์ RNA S. Ochoa ในปี 1959 ได้รับรางวัลโนเบล)
ไมโครโซม ไรโบโซม การแปล
ในช่วงกลางทศวรรษ 1950 เชื่อกันว่าไมโครโซมเป็นศูนย์กลางของการสังเคราะห์โปรตีนในเซลล์ คำว่า microsomes ถูกนำมาใช้ครั้งแรกในปี 1949 โดย A. Claude เพื่ออ้างถึงเศษส่วนของแกรนูลขนาดเล็ก ต่อมาปรากฎว่าไม่ใช่เศษส่วนของไมโครโซมทั้งหมดที่ประกอบด้วยเยื่อหุ้มและแกรนูล แต่มีเพียงอนุภาคไรโบนิวคลีโอโปรตีนขนาดเล็กเท่านั้นที่มีหน้าที่ในการสังเคราะห์โปรตีน อนุภาคเหล่านี้ในปี 1958 ถูกเรียกว่าไรโบโซมโดยอาร์. โรเบิร์ตส์
การศึกษาคลาสสิกของไรโบโซมของแบคทีเรียดำเนินการโดย A. Tisier และ J. Watson ในปี 1958-1959 ไรโบโซมของแบคทีเรียมีขนาดเล็กกว่าพืชและสัตว์ J. Littleton (1960), M. Clark (1964) และ E. N. Svetailo (1966) แสดงให้เห็นว่าไรโบโซมของคลอโรพลาสต์ของพืชชั้นสูงและไมโตคอนเดรียอยู่ในประเภทของแบคทีเรีย A. Tisier และคณะอื่นๆ (1958) พบว่าไรโบโซมแยกตัวออกเป็นสองหน่วยย่อยที่ไม่เท่ากันซึ่งมีโมเลกุล RNA หนึ่งโมเลกุลต่อหน่วย ในช่วงปลายยุค 50 เชื่อกันว่าโมเลกุล RNA ของไรโบโซมแต่ละโมเลกุลประกอบด้วยชิ้นส่วนสั้น ๆ หลายชิ้น อย่างไรก็ตาม AS Spirin ในปี 1960 เป็นครั้งแรกที่แสดงให้เห็นว่า RNA ในอนุภาคย่อยถูกแสดงด้วยโมเลกุลที่ต่อเนื่องกัน ด. วอลเลอร์ (1960) การแยกโปรตีนไรโบโซมโดยใช้สตาร์ชเจลอิเล็กโตรโฟรีซิส พบว่ามีความแตกต่างกันอย่างมาก ในตอนแรก หลายคนสงสัยในข้อมูลของวอลเลอร์ เนื่องจากดูเหมือนว่าโปรตีนไรโบโซมควรจะเป็นเนื้อเดียวกันอย่างเคร่งครัด เช่น โปรตีน TMV ในปัจจุบัน จากผลการวิจัยของ D. Waller, R. Trout, P. Traub และนักชีวเคมีอื่นๆ เป็นที่ทราบกันดีอยู่แล้วว่าองค์ประกอบของอนุภาคไรโบโซมที่เกิดขึ้นจริงประกอบด้วยโปรตีนมากกว่า 50 ชนิดที่มีโครงสร้างแตกต่างกันโดยสิ้นเชิง AS Spirin ในปีพ.ศ. 2506 เป็นรายแรกที่เปิดเผยอนุภาคย่อยของไรโบโซมและแสดงให้เห็นว่าไรโบโซมเป็นเส้นใยไรโบนิวคลีโอโปรตีนที่บิดเป็นเกลียวอย่างแน่นหนา ซึ่งสามารถกางออกได้ภายใต้สภาวะบางประการ ในปี 2510 - 2511 M. Nomura ได้สร้างหน่วยย่อยที่ออกฤทธิ์ทางชีวภาพขึ้นมาใหม่อย่างสมบูรณ์จากไรโบโซม RNA และโปรตีน และยังได้รับไรโบโซมซึ่งโปรตีนและอาร์เอ็นเอเป็นของจุลินทรีย์ต่างๆ
บทบาทของไรโบโซมอาร์เอ็นเอยังไม่ชัดเจน สันนิษฐานว่าเป็นเมทริกซ์จำเพาะที่มีลักษณะเฉพาะซึ่งในระหว่างการก่อตัวของอนุภาคไรโบโซม โปรตีนไรโบโซมจำนวนมากแต่ละตัวจะพบตำแหน่งที่กำหนดไว้อย่างเคร่งครัด (AS Spirin, 1968)
A. Rich (1962) ค้นพบการรวมตัวของไรโบโซมหลายตัวที่เชื่อมต่อกันด้วยเส้นใยของ mRNA คอมเพล็กซ์เหล่านี้เรียกว่าโพลีโซม การค้นพบโพลิโซมทำให้ริชและวัตสัน (1963) เสนอแนะว่าการสังเคราะห์สายโซ่โพลีเปปไทด์เกิดขึ้นบนไรโบโซม ซึ่งตามเดิม เคลื่อนที่ไปตามสายโซ่ mRNA ขณะที่ไรโบโซมเคลื่อนที่ไปตามสายโซ่ mRNA ข้อมูลจะถูกอ่านออกมาในอนุภาคและสายโปรตีนโพลีเปปไทด์จะถูกสร้างขึ้น และไรโบโซมใหม่จะเกาะติดกับส่วนปลายอ่านที่ปล่อยออกมาของ mRNA สลับกัน จากข้อมูลของ Rich และ Watson ตามมาว่าความสำคัญของโพลีโซมในเซลล์อยู่ที่การผลิตโปรตีนจำนวนมากโดยการอ่านเมทริกซ์อย่างต่อเนื่องโดยไรโบโซมหลายตัวในคราวเดียว
จากผลการวิจัยของ M. Nirenberg, S. Ochoa, F. Lipman, G. Korana และคณะอื่นๆ ในปี 1963 - 1970 เป็นที่ทราบกันว่าร่วมกับ mRNA, ไรโบโซม, ATP และ aminoacyl-tRNA ปัจจัยต่าง ๆ จำนวนมากมีส่วนร่วมในกระบวนการแปล และกระบวนการแปลนั้นสามารถแบ่งออกเป็นสามขั้นตอนตามเงื่อนไข - การเริ่มต้น การแปลเอง และการสิ้นสุด
การเริ่มต้นการแปลหมายถึงการสังเคราะห์พันธะเปปไทด์แรกในไรโบโซมที่ซับซ้อน - แม่แบบโพลีนิวคลีโอไทด์ - aminoacyl-tRNA การออกฤทธิ์เริ่มต้นดังกล่าวไม่ได้ครอบครองโดย aminoacyl-tRNA ใดๆ แต่โดย formylmethionyl-tRNA สารนี้ถูกแยกออกครั้งแรกในปี 2507 โดย F. Senger และ K. Marker S. Bretcher และ K. Marker (1966) แสดงให้เห็นว่าหน้าที่เริ่มต้นของ formylmethionyl-tRNA นั้นเกิดจากการที่มันสัมพันธ์กันเพิ่มขึ้นสำหรับจุดศูนย์กลางเปปทิดิลของไรโบโซม สำหรับการเริ่มต้นการแปล ปัจจัยการเริ่มโปรตีนบางอย่างก็มีความสำคัญอย่างยิ่งเช่นกัน ซึ่งแยกได้ในห้องทดลองของ S. Ochoa, F. Gro และศูนย์วิจัยอื่นๆ หลังจากการก่อรูปพันธะเปปไทด์แรกในไรโบโซม การแปลเองก็เริ่มต้น กล่าวคือ การเติมเรซิดิวของอะมิโนอะซิลตามลำดับไปยังปลาย C ของโพลีเปปไทด์ รายละเอียดมากมายของกระบวนการแปลได้รับการศึกษาโดย K. Monroe และ J. Bishop (อังกฤษ), I. Rykhlik และ F. Shorm (Czechoslovakia), F. Lipman, M. Bretcher, W. Gilbert (USA) และนักวิจัยคนอื่นๆ ในปี 1968 A. S. Spirin ได้เสนอสมมติฐานดั้งเดิมเพื่ออธิบายกลไกของไรโบโซม กลไกขับเคลื่อนที่ทำให้แน่ใจได้ว่าการเคลื่อนที่เชิงพื้นที่ทั้งหมดของ tRNA และ mRNA ระหว่างการแปลคือการเปิดและปิดเป็นระยะๆ ของอนุภาคย่อยไรโบโซม การสิ้นสุดการแปลถูกเข้ารหัสในเมทริกซ์ที่อ่านได้ซึ่งมีโคดอนการสิ้นสุด ดังที่แสดงโดย S. Brenner (1965 - 1967) แฝดสาม UAA, UAG และ UGA เป็น codon M. Capecci (1967) ยังระบุปัจจัยการยุติโปรตีนพิเศษอีกด้วย AS Spirin และ LP Gavrilova อธิบายการสังเคราะห์โปรตีนที่เรียกว่า "ไม่ใช่เอนไซม์" ในไรโบโซม (1972 - 1975) โดยไม่มีส่วนร่วมของปัจจัยโปรตีน การค้นพบนี้มีความสำคัญต่อการทำความเข้าใจที่มาและวิวัฒนาการของการสังเคราะห์โปรตีน
ระเบียบของยีนและการทำงานของโปรตีน
หลังจากปัญหาความจำเพาะของการสังเคราะห์โปรตีน ปัญหาของการควบคุมการสังเคราะห์โปรตีนหรือสิ่งที่เหมือนกันคือการควบคุมกิจกรรมของยีน กลับกลายเป็นว่าเกิดขึ้นที่แรกในอณูชีววิทยา
ความไม่เท่าเทียมกันในการทำงานของเซลล์และการกดขี่และการกระตุ้นยีนที่เกี่ยวข้องได้ดึงดูดความสนใจของนักพันธุศาสตร์มาช้านาน แต่กลไกที่แท้จริงในการควบคุมกิจกรรมของยีนยังไม่ทราบจนกระทั่งเมื่อไม่นานมานี้
ความพยายามครั้งแรกในการอธิบายกิจกรรมการควบคุมยีนเกี่ยวข้องกับการศึกษาโปรตีนฮิสโตน แม้แต่คู่สมรสของ Steadman * ในช่วงต้นยุค 40 ของศตวรรษที่ XX แนะนำว่าเป็นฮิสโตนที่มีบทบาทสำคัญในปรากฏการณ์นี้ ต่อจากนั้น พวกเขาได้รับข้อมูลที่ชัดเจนเป็นครั้งแรกเกี่ยวกับความแตกต่างในลักษณะทางเคมีของโปรตีนฮิสโตน ในปัจจุบันข้อเท็จจริงที่เป็นพยานสนับสนุนสมมติฐานนี้มีจำนวนเพิ่มขึ้นทุกปี
* (อี. สเตดแมน, อี. สเตดแมน. โปรตีนพื้นฐานของนิวเคลียสของเซลล์.- ปรัชญา. ทรานส์ รอย. ซ. ลอนดอน 2494 วี. 235, 565 - 595.)
ในเวลาเดียวกัน ข้อมูลจำนวนมากขึ้นกำลังสะสมอยู่ ซึ่งบ่งชี้ว่าการควบคุมการทำงานของยีนเป็นกระบวนการที่ซับซ้อนกว่าการโต้ตอบอย่างง่ายของส่วนของยีนกับโมเลกุลโปรตีนฮิสโตน ในปี 1960 - 1962 ในห้องปฏิบัติการของ R. B. Khesin-Lurie พบว่ายีน phage เริ่มอ่านไม่พร้อมกัน: ยีน T2 phage สามารถแบ่งออกเป็นยีนแรก ๆ ซึ่งการทำงานเกิดขึ้นในนาทีแรกของการติดเชื้อของเซลล์แบคทีเรียและ สายซึ่งเริ่มสังเคราะห์ mRNA หลังจากเสร็จสิ้นการทำงานของยีนต้น
ในปีพ.ศ. 2504 นักชีวเคมีชาวฝรั่งเศส F. Jacob และ J. Monod ได้เสนอโครงการควบคุมการทำงานของยีน ซึ่งมีบทบาทสำคัญในการทำความเข้าใจกลไกการกำกับดูแลของเซลล์โดยทั่วไป ตามโครงการของจาค็อบและโมโนด นอกเหนือจากยีนที่มีโครงสร้าง (ข้อมูล) แล้ว DNA ยังประกอบด้วยตัวควบคุมยีนและตัวดำเนินการยีนด้วย ยีนควบคุมเข้ารหัสการสังเคราะห์สารเฉพาะ - สารยับยั้ง ซึ่งสามารถยึดติดกับทั้งตัวเหนี่ยวนำและยีนตัวดำเนินการ ยีนตัวดำเนินการเชื่อมโยงกับยีนโครงสร้าง ในขณะที่ยีนควบคุมอยู่ห่างจากพวกมัน หากไม่มีตัวกระตุ้นในสิ่งแวดล้อมเช่นแลคโตสสารยับยั้งที่สังเคราะห์โดยยีนควบคุมจะจับกับยีนตัวดำเนินการและปิดกั้นการทำงานของโอเปอเรเตอร์ทั้งหมด (บล็อกของยีนที่มีโครงสร้างพร้อมกับโอเปอเรเตอร์ ที่ควบคุมพวกเขา) การก่อตัวของเอนไซม์จะไม่เกิดขึ้นภายใต้สภาวะเหล่านี้ หากตัวเหนี่ยวนำ (แลคโตส) ปรากฏขึ้นในตัวกลาง ผลิตภัณฑ์ของยีนควบคุม ตัวยับยั้งจะจับกับแลคโตสและดึงบล็อกออกจากยีนตัวดำเนินการ ในกรณีนี้ การทำงานของยีนโครงสร้างที่เข้ารหัสการสังเคราะห์เอ็นไซม์เป็นไปได้ และเอ็นไซม์ (แลคโตส) จะปรากฏในตัวกลาง
จากคำกล่าวของยาโคบและโมโนด รูปแบบการควบคุมนี้ใช้ได้กับเอ็นไซม์ที่ปรับตัวได้ทั้งหมด และสามารถเกิดขึ้นได้ทั้งในระหว่างการกดขี่ เมื่อการก่อตัวของเอ็นไซม์ถูกกดโดยผลิตภัณฑ์ปฏิกิริยาที่มากเกินไป และในระหว่างการเหนี่ยวนำ เมื่อการแนะนำของซับสเตรตทำให้เกิด การสังเคราะห์เอนไซม์ สำหรับการศึกษาการควบคุมกิจกรรมของยีน ยาโคบและโมโนดได้รับรางวัลโนเบลในปี พ.ศ. 2508
ในขั้นต้น แผนการนี้ดูไกลเกินเอื้อม อย่างไรก็ตาม ภายหลังปรากฎว่าการควบคุมยีนตามหลักการนี้ไม่ได้เกิดขึ้นเฉพาะในแบคทีเรียเท่านั้น แต่ยังเกิดขึ้นในสิ่งมีชีวิตอื่นๆ ด้วย
ตั้งแต่ปี 1960 สถานที่ที่โดดเด่นในอณูชีววิทยาได้ถูกครอบครองโดยการศึกษาเกี่ยวกับการจัดระเบียบของจีโนมและโครงสร้างของโครมาตินในสิ่งมีชีวิตที่มียูคาริโอต (J. Bonner, R. Britten, W. Olfrey, P. Walker, Yu. S. Chentsov , I. B. Zbarsky และอื่น ๆ . ) และกฎระเบียบของการถอดความ (A. Mirsky, G. P. Georgiev, M. Bernstiel, D. Goll, R. Tsanev, R. I. Salganik) เป็นเวลานานที่ธรรมชาติของผู้กดขี่ยังไม่ทราบและเป็นที่ถกเถียงกัน ในปี 1968 M. Ptashne (USA) แสดงให้เห็นว่าโปรตีนเป็นตัวยับยั้ง เขาแยกมันออกจากห้องทดลองของเจ. วัตสัน และพบว่าสารยับยั้งมีความเกี่ยวข้องกับตัวเหนี่ยวนำ (แลคโตส) และในขณะเดียวกัน "รับรู้" ยีนตัวดำเนินการของครั่ง โอเปร่า และผูกมัดกับมันโดยเฉพาะ
ในช่วง 5-7 ปีที่ผ่านมา ได้รับข้อมูลเกี่ยวกับการมีอยู่ของเซลล์ควบคุมอื่นของกิจกรรมของยีน - โปรโมเตอร์ ปรากฎว่าในพื้นที่ใกล้เคียงของไซต์ผู้ปฏิบัติงานซึ่งผลิตภัณฑ์สังเคราะห์บนตัวควบคุมยีน - สารโปรตีนของตัวยับยั้งถูกแนบมีไซต์อื่นซึ่งควรนำมาประกอบกับสมาชิกของระบบการกำกับดูแล ของกิจกรรมของยีน โมเลกุลโปรตีนของเอนไซม์ RNA polymerase ติดอยู่ที่ไซต์นี้ ในภูมิภาคโปรโมเตอร์ จะต้องรับรู้ร่วมกันของลำดับนิวคลีโอไทด์ที่ไม่ซ้ำกันใน DNA และการกำหนดค่าเฉพาะของโปรตีน RNA polymerase การดำเนินการตามกระบวนการอ่านข้อมูลทางพันธุกรรมด้วยลำดับของยีนของโอเปอเรเตอร์ที่อยู่ติดกับโปรโมเตอร์จะขึ้นอยู่กับประสิทธิภาพการจดจำ
นอกจากโครงร่างที่อธิบายโดยยาโคบและโมโนดแล้ว ยังมีกลไกอื่นๆ ของการควบคุมยีนในเซลล์อีกด้วย F. Jacob และ S. Brenner (1963) กำหนดว่าการควบคุมการจำลองแบบ DNA ของแบคทีเรียถูกควบคุมโดยเยื่อหุ้มเซลล์ด้วยวิธีใดวิธีหนึ่ง การทดลองของยาโคบ (1954) เกี่ยวกับการชักนำให้เกิดการเผยพระวจนะต่างๆ แสดงให้เห็นอย่างน่าเชื่อถือว่าภายใต้อิทธิพลของปัจจัยการกลายพันธุ์ต่างๆ ในเซลล์ของแบคทีเรียไลโซเจนิค การจำลองแบบคัดเลือกของยีนการเผยพระวจนะเริ่มต้นขึ้น และการจำลองแบบของจีโนมเจ้าบ้านถูกปิดกั้น ในปี 1970 เอฟ. เบลล์รายงานว่าโมเลกุลดีเอ็นเอขนาดเล็กสามารถส่งผ่านจากนิวเคลียสไปยังไซโทพลาซึมและถ่ายทอดที่นั่นได้
ดังนั้น กิจกรรมของยีนสามารถควบคุมได้ที่ระดับการจำลองแบบ การถอดความ และการแปล
มีความก้าวหน้าอย่างมากในการศึกษากฎระเบียบไม่เพียงแต่การสังเคราะห์เอนไซม์เท่านั้น แต่ยังรวมถึงกิจกรรมของเอนไซม์ด้วย A. Novik และ L. Szilard ชี้ให้เห็นปรากฏการณ์ของการควบคุมการทำงานของเอนไซม์ในเซลล์ในช่วงปี 1950 G. Umbarger (1956) พบว่าในเซลล์มีวิธีการที่มีเหตุผลมากในการยับยั้งการทำงานของเอนไซม์โดยผลิตภัณฑ์สุดท้ายของห่วงโซ่การป้อนกลับของปฏิกิริยา ตามที่กำหนดโดย J. Monod, J. Change, F. Jacob, A. Purdy และนักวิจัยคนอื่น ๆ (1956 - 1960) การควบคุมการทำงานของเอนไซม์สามารถทำได้ตามหลักการ allosteric เอ็นไซม์หรือหน่วยย่อยหนึ่งของเอ็นไซม์ นอกเหนือจากค่าสัมพรรคภาพสำหรับซับสเตรตแล้ว ยังมีสัมพรรคภาพกับหนึ่งในผลิตภัณฑ์ของห่วงโซ่ปฏิกิริยา ภายใต้อิทธิพลของผลิตภัณฑ์สัญญาณดังกล่าว เอ็นไซม์จะเปลี่ยนโครงสร้างในลักษณะที่สูญเสียกิจกรรม เป็นผลให้ห่วงโซ่ปฏิกิริยาเอนไซม์ทั้งหมดถูกปิดในตอนเริ่มต้น D. Wieman และ R. Woodward (1952; ผู้ได้รับรางวัลโนเบล, 1965) ชี้ให้เห็นถึงบทบาทที่สำคัญของการเปลี่ยนแปลงโครงสร้างของโปรตีนในปฏิกิริยาของเอนไซม์ และในแง่หนึ่ง การมีอยู่ของผล allosteric
โครงสร้างและหน้าที่ของโปรตีน
อันเป็นผลมาจากผลงานของ T. Osborn, G. Hofmeister, A. Gurber, F. Schulz และอีกหลายคนในช่วงปลายศตวรรษที่ 19 โปรตีนจากสัตว์และพืชหลายชนิดได้รับในรูปแบบผลึก ในเวลาเดียวกัน น้ำหนักโมเลกุลของโปรตีนบางชนิดถูกกำหนดโดยใช้วิธีการทางกายภาพต่างๆ ดังนั้นในปี 1891 A. Sabaneev และ N. Alexandrov รายงานว่าน้ำหนักโมเลกุลของ ovalbumin คือ 14,000; ในปี 1905 E. Reid พบว่าน้ำหนักโมเลกุลของเฮโมโกลบินเท่ากับ 48,000 โครงสร้างโพลีเมอร์ของโปรตีนถูกค้นพบในปี 1871 โดย G. Glasivetz และ D. Gaberman แนวคิดของพันธะเปปไทด์ของกรดอะมิโนแต่ละตัวที่ตกค้างในโปรตีนถูกนำเสนอโดย T. Curtius (1883) ทำงานเกี่ยวกับการควบแน่นทางเคมีของกรดอะมิโน (E. Schaal, 1871; G. Schiff, 1897; L. Balbiano and D. Traschiatti, 1900) และการสังเคราะห์เฮเทอโรโพลีเปปไทด์ (E. Fisher, 1902 - 1907, Nobel Prize, 1902) นำไปสู่การพัฒนาหลักการพื้นฐานของโครงสร้างทางเคมีของโปรตีน
เอนไซม์ที่เป็นผลึกตัวแรก (urease) ได้รับในปี 1926 โดย J. Sumner (รางวัลโนเบล, 1946) และในปี 1930 J. Northrop (รางวัลโนเบล, 1946) ได้รับผลึกเปปซิน หลังจากการทำงานเหล่านี้ เป็นที่ชัดเจนว่าเอ็นไซม์มีลักษณะเป็นโปรตีน ในปี 1940 M. Kunits ได้แยกผลึก RNase ภายในปี 1958 มีเอนไซม์ที่เป็นผลึกมากกว่า 100 ชนิดและเอ็นไซม์ที่ไม่ใช่ผลึกมากกว่า 500 ชนิดเป็นที่ทราบกันดีอยู่แล้ว การเตรียมโปรตีนแต่ละชนิดที่มีความบริสุทธิ์สูงมีส่วนช่วยในการถอดรหัสโครงสร้างหลักและการจัดโครงสร้างโมเลกุลขนาดใหญ่
สิ่งที่สำคัญอย่างยิ่งสำหรับการพัฒนาของอณูชีววิทยาโดยทั่วไปและโดยเฉพาะอย่างยิ่งพันธุศาสตร์ของมนุษย์คือการค้นพบโดย L. Pauling (1940) ของฮีโมโกลบิน S ที่ผิดปกติซึ่งแยกได้จากเม็ดเลือดแดงของผู้ที่มีโรคทางพันธุกรรมรุนแรง โรคโลหิตจางชนิดเคียว ในปี พ.ศ. 2498 - 2500 W. Ingram ใช้วิธีการ "ลายนิ้วมือ" ที่พัฒนาโดย F. Sanger (จุดที่เกิดจากเปปไทด์แต่ละตัวระหว่างโครมาโตกราฟีบนกระดาษ) เพื่อวิเคราะห์ผลิตภัณฑ์ของการไฮโดรไลซิสของเฮโมโกลบิน S ด้วยด่างและทริปซิน ในปี 1961 Ingram รายงานว่าเฮโมโกลบิน S แตกต่างจากเฮโมโกลบินปกติในลักษณะของกรดอะมิโนตัวเดียวเท่านั้น: ในเฮโมโกลบินปกติ กรดกลูตามิกตกค้างอยู่ในตำแหน่งที่เจ็ดของสายโซ่ และในเฮโมโกลบิน S ซึ่งเป็นสารตกค้างของวาลีน ดังนั้นสมมติฐานของ Pauling (1949) ที่ว่าโรคโลหิตจางชนิดเคียวเป็นโรคที่มีลักษณะโมเลกุลได้รับการยืนยันอย่างสมบูรณ์ การเปลี่ยนแปลงที่สืบทอดมาของกรดอะมิโนเพียงตัวเดียวในแต่ละครึ่งของโมเลกุลขนาดใหญ่ของเฮโมโกลบินทำให้ฮีโมโกลบินสูญเสียความสามารถในการละลายได้ง่ายที่ความเข้มข้นของออกซิเจนต่ำและเริ่มตกผลึก ซึ่งนำไปสู่การหยุดชะงักของโครงสร้างเซลล์ การศึกษาเหล่านี้แสดงให้เห็นอย่างชัดเจนว่าโครงสร้างของโปรตีนเป็นลำดับกรดอะมิโนที่กำหนดไว้อย่างเคร่งครัดซึ่งเข้ารหัสในจีโนม ผลงานของ K. Anfinsen (1951) พิสูจน์ให้เห็นถึงความสำคัญพิเศษของโครงสร้างหลักของโปรตีนในการก่อตัวของโครงสร้างทางชีวภาพที่ไม่เหมือนใครของโมเลกุลขนาดใหญ่ Anfinsen แสดงให้เห็นว่าโครงสร้างมหภาคที่ใช้งานทางชีวภาพของไรโบนิวคลีเอสในตับอ่อนซึ่งสูญเสียไปอันเป็นผลมาจากการฟื้นฟูนั้นถูกกำหนดไว้ล่วงหน้าโดยลำดับกรดอะมิโนและสามารถปรากฏขึ้นอีกครั้งได้เองตามธรรมชาติในระหว่างการออกซิเดชันของกลุ่ม SH ของซิสเทอีนที่ตกค้างด้วยการก่อตัวของพันธะซัลไฟด์อย่างเคร่งครัด ตำแหน่งที่กำหนดของสายโซ่เปปไทด์ของเอนไซม์
จนถึงปัจจุบันมีการศึกษากลไกการออกฤทธิ์ของเอ็นไซม์จำนวนมากอย่างละเอียดและได้กำหนดโครงสร้างของโปรตีนหลายชนิด
ในปีพ.ศ. 2496 เอฟ. แซงเจอร์ได้กำหนดลำดับกรดอะมิโนของอินซูลิน : โปรตีนนี้ประกอบด้วยสายโซ่โพลีเปปไทด์สองสายที่เชื่อมต่อกันด้วยตัวเชื่อมขวางไดซัลไฟด์สองสาย สายหนึ่งมีกรดอะมิโนเพียง 21 ตัว ในขณะที่อีกสายมี 30 เรซิดิว แซงเจอร์ใช้เวลาประมาณ 10 ปีในการถอดรหัสโครงสร้างของโปรตีนที่ค่อนข้างง่ายนี้ ในปี 1958 เขาได้รับรางวัลโนเบลสำหรับงานวิจัยที่โดดเด่นนี้ หลังจากการสร้างโดย V. Stein และ S. Moore (1957) ของเครื่องวิเคราะห์กรดอะมิโนอัตโนมัติ การระบุผลิตภัณฑ์ของการไฮโดรไลซิสบางส่วนของโปรตีนเร่งขึ้นอย่างมีนัยสำคัญ ในปี 1960 สไตน์และมัวร์ได้รายงานไปแล้วว่า ว่าพวกเขาสามารถกำหนดลำดับของไรโบนิวคลีเอสซึ่งเป็นสายโซ่เปปไทด์ซึ่งมีกรดอะมิโน 124 ตัวแสดงแทน ในปีเดียวกันนั้น ในห้องปฏิบัติการของ G. Schramm ใน Tübingen (เยอรมนี) F. Anderer และคนอื่นๆ ได้กำหนดลำดับกรดอะมิโนในโปรตีน TMV จากนั้นกำหนดลำดับกรดอะมิโนใน myoglobin (A. Edmunson) และ α- และ β-chains ของเฮโมโกลบินของมนุษย์ (G. Braunitzer, E. Schroeder เป็นต้น) lysozyme จากโปรตีนไข่ (J. Jollet, D. Keyfield) . ในปี 1963 F. Shorm และ B. Keil (Czechoslovakia) ได้สร้างลำดับกรดอะมิโนในโมเลกุลของ chymotrypsinogen ในปีเดียวกันนั้น ได้มีการกำหนดลำดับกรดอะมิโนของทริปซิโนเจน (F. Shorm, D. Walsh) ในปี 1965 K. Takahashi ได้ก่อตั้งโครงสร้างหลักของไรโบนิวคลีเอส T1 จากนั้นจึงกำหนดลำดับกรดอะมิโนสำหรับโปรตีนอีกหลายตัว
ดังที่ทราบกันดีว่าข้อพิสูจน์ขั้นสุดท้ายของความถูกต้องของคำจำกัดความของโครงสร้างเฉพาะคือการสังเคราะห์ ในปี 1969 R. Merifield (สหรัฐอเมริกา) เป็นคนแรกที่ทำการสังเคราะห์ทางเคมีของไรโบนิวคลีเอสในตับอ่อน ด้วยความช่วยเหลือของวิธีการสังเคราะห์ของเขาบนตัวพาเฟสของแข็ง Merifield ได้เพิ่มกรดอะมิโนหนึ่งตัวหลังจากนั้นอีกตัวหนึ่งไปยังสายโซ่ตามลำดับที่อธิบายโดยสไตน์และมัวร์ เป็นผลให้เขาได้รับโปรตีนที่มีคุณภาพเหมือนกันกับไรโบนิวคลีเอสตับอ่อน A สำหรับการค้นพบโครงสร้างของไรโบนิวคลีเอส V. Stein, S. Moore และ K. Anfinsen ได้รับรางวัลโนเบลในปี 1972 การสังเคราะห์โปรตีนตามธรรมชาตินี้เปิดโอกาสที่ดี โดยชี้ให้เห็นถึงความเป็นไปได้ในการสร้างโปรตีนใดๆ ตามลำดับที่วางแผนไว้ล่วงหน้า
จากการศึกษาเอ็กซ์เรย์โดย W. Astbury (1933) พบว่าสายโซ่เปปไทด์ของโมเลกุลโปรตีนบิดหรือซ้อนกันในลักษณะที่กำหนดไว้อย่างเข้มงวด นับตั้งแต่นั้นมา ผู้เขียนหลายคนได้แสดงสมมติฐานต่างๆ เกี่ยวกับวิธีการพับสายโปรตีน แต่จนถึงปี 1951 แบบจำลองทั้งหมดยังคงเป็นโครงสร้างการเก็งกำไรที่ไม่สอดคล้องกับข้อมูลการทดลอง ในปี 1951 L. Pauling และ R. Corey ได้ตีพิมพ์ผลงานที่ยอดเยี่ยมหลายชุด ซึ่งในที่สุดทฤษฎีของโครงสร้างรองของโปรตีน ทฤษฎีของ α-helix ก็ได้ถูกสร้างขึ้น นอกจากนี้ยังเป็นที่ทราบกันดีว่าโปรตีนยังมีโครงสร้างตติยภูมิอีกด้วย: α-helix ของสายเปปไทด์สามารถพับในลักษณะที่แน่นอนทำให้เกิดโครงสร้างที่ค่อนข้างกะทัดรัด
ในปี 1957 J. Kendrew และผู้ทำงานร่วมกันได้เสนอแบบจำลองสามมิติของโครงสร้างของ myoglobin แบบจำลองนี้ได้รับการขัดเกลามาเป็นเวลาหลายปี จนกระทั่งงานสุดท้ายปรากฏขึ้นในปี 2504 โดยมีลักษณะเฉพาะของโครงสร้างเชิงพื้นที่ของโปรตีนนี้ ในปี 1959 M. Perutz และเพื่อนร่วมงานได้ก่อตั้งโครงสร้างสามมิติของเฮโมโกลบิน นักวิจัยใช้เวลามากกว่า 20 ปีในงานนี้ (Perutz ได้รับรังสีเอกซ์ครั้งแรกของเฮโมโกลบินในปี 2480) เนื่องจากโมเลกุลของเฮโมโกลบินประกอบด้วยหน่วยย่อยสี่หน่วย ซึ่งได้ถอดรหัสการจัดระเบียบของมัน เปรุตซ์จึงอธิบายโครงสร้างควอเทอร์นารีของโปรตีนก่อน สำหรับงานของพวกเขาในการกำหนดโครงสร้างสามมิติของโปรตีน Kendrew และ Perutz ได้รับรางวัลโนเบลในปี 2505
การสร้างแบบจำลองเชิงพื้นที่ของโครงสร้างของเฮโมโกลบินโดย Perutz ได้รับอนุญาต เพื่อให้เข้าใจกลไกการทำงานของโปรตีนมากขึ้น ซึ่งอย่างที่ทราบกันดีอยู่แล้วว่าทำหน้าที่ขนส่งออกซิเจนในเซลล์สัตว์ ย้อนกลับไปในปี 2480 F. Gaurowitz ได้ข้อสรุปว่าปฏิกิริยาของเฮโมโกลบินกับออกซิเจนอากาศควรมาพร้อมกับการเปลี่ยนแปลงโครงสร้างของโปรตีน ในทศวรรษที่ 1960 Perutz และเพื่อนร่วมงานค้นพบการเปลี่ยนแปลงที่เห็นได้ชัดเจนในสายโซ่เฮโมโกลบินหลังจากการออกซิเดชันซึ่งเกิดจากการเปลี่ยนของอะตอมของเหล็กอันเป็นผลมาจากการจับกับออกซิเจน บนพื้นฐานนี้มีการสร้างแนวคิดเกี่ยวกับ "การหายใจ" ของโมเลกุลโปรตีนขนาดใหญ่ขึ้น
ในปีพ.ศ. 2503 ดี. ฟิลลิปส์และผู้ทำงานร่วมกันได้เริ่มการศึกษาการเลี้ยวเบนของรังสีเอกซ์ของโมเลกุลไลโซไซม์ ภายในปี 1967 พวกมันสามารถสร้างรายละเอียดเกี่ยวกับการจัดระเบียบของโปรตีนนี้และการโลคัลไลเซชันของอะตอมแต่ละตัวในโมเลกุลของมันได้ไม่มากก็น้อย นอกจากนี้ ฟิลลิปส์ยังได้ค้นพบธรรมชาติของการเติมไลโซไซม์ลงในสารตั้งต้น (ไตรอะเซทิลกลูโคซามีน) ทำให้สามารถสร้างกลไกของเอนไซม์นี้ขึ้นมาใหม่ได้ ดังนั้น ความรู้เกี่ยวกับโครงสร้างหลักและการจัดระเบียบโมเลกุลขนาดใหญ่จึงทำให้ไม่เพียงแต่สร้างธรรมชาติของศูนย์กลางการทำงานของเอนไซม์จำนวนมากเท่านั้น แต่ยังเผยให้เห็นกลไกการทำงานของโมเลกุลขนาดใหญ่เหล่านี้อย่างเต็มที่
การใช้กล้องจุลทรรศน์อิเล็กตรอนแบบส่องกราดช่วยเผยให้เห็นหลักการของการจัดระเบียบโมเลกุลขนาดใหญ่ของการสร้างโปรตีนที่ซับซ้อน เช่น คอลลาเจน ไฟบริโนเจน เส้นใยกล้ามเนื้อหดตัว เป็นต้น ในช่วงปลายทศวรรษ 1950 มีการเสนอแบบจำลองของอุปกรณ์หดตัวของกล้ามเนื้อ สิ่งสำคัญอย่างยิ่งในการทำความเข้าใจกลไกการหดตัวของกล้ามเนื้อคือการค้นพบโดย V. A. Engelgardt และ M. N. Lyubimova (1939) ของกิจกรรม ATPase ของ myosin ซึ่งหมายความว่าการหดตัวของกล้ามเนื้อขึ้นอยู่กับการเปลี่ยนแปลงคุณสมบัติทางเคมีกายภาพและการจัดระเบียบโมเลกุลขนาดใหญ่ของโปรตีนหดตัวภายใต้อิทธิพลของกรดอะดีโนซีนไตรฟอสฟอริก (ดูบทที่ 11)
การวิจัยทางไวรัสวิทยามีความสำคัญต่อการทำความเข้าใจหลักการของการประกอบโครงสร้างทางชีววิทยา (ดูบทที่ 25)
ปัญหาที่ไม่ได้รับการแก้ไข
ความก้าวหน้าหลักในอณูชีววิทยาสมัยใหม่เกิดขึ้นจากการศึกษากรดนิวคลีอิกเป็นหลัก อย่างไรก็ตามแม้ในพื้นที่นี้จะห่างไกลจากปัญหาทั้งหมดได้รับการแก้ไข โดยเฉพาะอย่างยิ่งจะต้องใช้ความพยายามอย่างมากในการถอดรหัสลำดับนิวคลีโอไทด์ทั้งหมดของจีโนม ในทางกลับกัน ปัญหานี้เชื่อมโยงกับปัญหาความแตกต่างของ DNA อย่างแยกไม่ออก และจำเป็นต้องมีการพัฒนาวิธีการขั้นสูงใหม่สำหรับการแยกส่วนและการแยกโมเลกุลแต่ละส่วนออกจากสารพันธุกรรมทั้งหมดของเซลล์
จนถึงปัจจุบัน ความพยายามส่วนใหญ่มุ่งเน้นไปที่การศึกษาโปรตีนและกรดนิวคลีอิกแยกจากกัน ในเซลล์ ไบโอโพลีเมอร์เหล่านี้เชื่อมโยงกันอย่างแยกไม่ออกและทำงานเป็นส่วนใหญ่ในรูปของนิวคลีโอโปรตีน ดังนั้นความจำเป็นในการศึกษาปฏิสัมพันธ์ของโปรตีนและกรดนิวคลีอิกจึงรุนแรงขึ้นเป็นพิเศษ ปัญหาในการรับรู้กรดนิวคลีอิกบางส่วนโดยโปรตีนถูกกล่าวถึงในเบื้องต้น มีการกำหนดขั้นตอนในการศึกษาปฏิสัมพันธ์ดังกล่าวของพอลิเมอร์ชีวภาพเหล่านี้แล้ว โดยที่ไม่เข้าใจโครงสร้างและหน้าที่ของโครโมโซม ไรโบโซม และโครงสร้างอื่นๆ อย่างถ่องแท้ หากปราศจากสิ่งนี้ ก็ยังเป็นไปไม่ได้ที่จะเข้าใจกฎระเบียบของกิจกรรมของยีนและในที่สุดก็ถอดรหัสหลักการทำงานของกลไกการสังเคราะห์โปรตีน หลังจากงานของ Jacob และ Monod ข้อมูลใหม่บางส่วนปรากฏขึ้นเกี่ยวกับความสำคัญด้านกฎระเบียบของเมมเบรนในการสังเคราะห์วัสดุนิวเคลียร์ ทำให้เกิดปัญหาในการศึกษาบทบาทของเยื่อหุ้มเซลล์ในการควบคุมการจำลองดีเอ็นเออย่างลึกซึ้ง โดยทั่วไป ปัญหาของการควบคุมการทำงานของยีนและการทำงานของเซลล์โดยทั่วไปได้กลายเป็นปัญหาที่สำคัญที่สุดอย่างหนึ่งของอณูชีววิทยาสมัยใหม่
สถานะปัจจุบันของชีวฟิสิกส์
ในการเชื่อมต่ออย่างใกล้ชิดกับปัญหาของอณูชีววิทยา การพัฒนาชีวฟิสิกส์ดำเนินไป ด้านหนึ่งความสนใจในด้านชีววิทยาถูกกระตุ้นโดยความจำเป็นในการศึกษาผลกระทบของรังสีประเภทต่าง ๆ ที่มีต่อร่างกายอย่างครอบคลุมและในทางกลับกันโดยความต้องการศึกษาทางกายภาพและทางกายภาพ - พื้นฐานทางเคมีของปรากฏการณ์ชีวิตที่เกิดขึ้นในระดับโมเลกุล
การได้รับข้อมูลที่ถูกต้องเกี่ยวกับโครงสร้างโมเลกุลและกระบวนการที่เกิดขึ้นนั้นเป็นไปได้อันเป็นผลมาจากการใช้วิธีการทางกายภาพและเคมีที่ดีแบบใหม่ จากความสำเร็จของเคมีไฟฟ้า เป็นไปได้ที่จะปรับปรุงวิธีการวัดศักย์ไฟฟ้าชีวภาพโดยใช้อิเล็กโทรดคัดเลือกไอออน (G. Eisenman, B.P. Nikolsky, Khuri, 50-60s) อินฟราเรดสเปกโตรสโคปี (ด้วยการใช้อุปกรณ์เลเซอร์) เพิ่มมากขึ้นเรื่อยๆ ซึ่งทำให้สามารถศึกษาการเปลี่ยนแปลงโครงสร้างของโปรตีนได้ (I. Plotnikov, 1940) ข้อมูลที่มีค่ายังถูกจัดเตรียมโดยวิธีการเรโซแนนซ์อิเล็กตรอนแบบพาราแมกเนติก (E.K. Zavoisky, 1944) และวิธีการทางชีวเคมี (B. N. Tarusov et al., 1960) ซึ่งทำให้สามารถตัดสินการขนส่งอิเล็กตรอนในระหว่างกระบวนการออกซิเดชัน
ในช่วงทศวรรษ 1950 ชีวฟิสิกส์ได้รับตำแหน่งที่แข็งแกร่งแล้ว จำเป็นต้องฝึกอบรมผู้เชี่ยวชาญที่มีคุณสมบัติเหมาะสม หากในปี 1911 ในยุโรปมีเพียง University of Pécs ในฮังการีเท่านั้นที่มีประธานด้านชีวฟิสิกส์ จากนั้นในปี 1973 เก้าอี้ดังกล่าวจะมีอยู่ในมหาวิทยาลัยหลักๆ เกือบทั้งหมด
ในปีพ. ศ. 2503 ได้มีการจัดตั้งสมาคมนักชีวฟิสิกส์ระหว่างประเทศขึ้น ในเดือนสิงหาคม พ.ศ. 2504 การประชุมทางชีวฟิสิกส์ระหว่างประเทศครั้งแรกจัดขึ้นที่กรุงสตอกโฮล์ม การประชุมครั้งที่สองจัดขึ้นในปี 2508 ที่ปารีสครั้งที่สาม - ในปี 2512 ที่บอสตันครั้งที่สี่ในปี 2515 ที่กรุงมอสโก
ในชีวฟิสิกส์ มีความแตกต่างที่ชัดเจนระหว่างสองส่วนที่มีเนื้อหาต่างกัน - ชีวฟิสิกส์ระดับโมเลกุลและชีวฟิสิกส์ระดับเซลล์ ความแตกต่างนี้ยังได้รับการแสดงออกขององค์กร: มีการสร้างแผนกที่แยกจากกันของสองด้านของชีวฟิสิกส์ ที่มหาวิทยาลัยมอสโก ภาควิชาชีวฟิสิกส์แห่งแรกก่อตั้งขึ้นในปี 2496 ที่คณะชีววิทยาและดินศาสตร์ และอีกไม่นานภาควิชาชีวฟิสิกส์ก็ปรากฏตัวที่คณะฟิสิกส์ หน่วยงานต่างๆ ถูกจัดระเบียบตามหลักการเดียวกันในมหาวิทยาลัยอื่นๆ หลายแห่ง
ชีวฟิสิกส์ระดับโมเลกุล
ในช่วงไม่กี่ปีที่ผ่านมา ความเชื่อมโยงระหว่างชีวฟิสิกส์ระดับโมเลกุลและอณูชีววิทยามีความเข้มแข็งมากขึ้น และตอนนี้บางครั้งก็ยากที่จะระบุได้ว่าเส้นแบ่งระหว่างพวกมันอยู่ที่ใด ในการโจมตีปัญหาข้อมูลทางพันธุกรรมโดยทั่วไป ความร่วมมือดังกล่าวระหว่างชีวฟิสิกส์และอณูชีววิทยาเป็นสิ่งที่หลีกเลี่ยงไม่ได้
ทิศทางหลักในงานวิจัยคือการศึกษาฟิสิกส์ของกรดนิวคลีอิก - DNA และ RNA การใช้วิธีการข้างต้นและเหนือสิ่งอื่นใด การวิเคราะห์การเลี้ยวเบนของรังสีเอกซ์มีส่วนทำให้เกิดการถอดรหัสโครงสร้างโมเลกุลของกรดนิวคลีอิก ขณะนี้ การวิจัยอย่างเข้มข้นกำลังดำเนินการศึกษาพฤติกรรมของกรดเหล่านี้ในสารละลาย "ขดลวดเกลียว" ซึ่งศึกษาโดยการเปลี่ยนแปลงค่าความหนืด พารามิเตอร์ทางแสงและทางไฟฟ้า ในการศึกษากลไกการกลายพันธุ์ ได้มีการพัฒนาการศึกษาเพื่อศึกษาผลของการแผ่รังสีไอออไนซ์ต่อพฤติกรรมของกรดนิวคลีอิกในสารละลาย ตลอดจนผลกระทบของการแผ่รังสีต่อกรดนิวคลีอิกของไวรัสและฟาจ ผลของรังสีอัลตราไวโอเลต ซึ่งบางบริเวณสเปกตรัมเป็นที่ทราบกันว่ากรดนิวคลีอิกดูดซับได้ดี ต้องได้รับการวิเคราะห์อย่างครอบคลุม งานวิจัยประเภทนี้ส่วนใหญ่เกิดจากการตรวจหาอนุมูลอิสระของกรดนิวคลีอิกและโปรตีนโดยวิธีอิเล็กตรอนพาราแมกเนติกเรโซแนนซ์ ด้วยการใช้วิธีนี้ การเกิดขึ้นของทิศทางที่เป็นอิสระทั้งหมดสัมพันธ์กัน
ปัญหาในการเข้ารหัสข้อมูล DNA และ RNA และการส่งผ่านข้อมูลระหว่างการสังเคราะห์โปรตีนเป็นที่สนใจของชีวฟิสิกส์ระดับโมเลกุลมาช้านาน และนักฟิสิกส์ได้แสดงการพิจารณาบางอย่างซ้ำแล้วซ้ำเล่าในเรื่องนี้ (E. Schrödinger, G. Gamow) การถอดรหัสรหัสพันธุกรรมทำให้เกิดการศึกษาเชิงทฤษฎีและเชิงทดลองมากมายเกี่ยวกับโครงสร้างของเกลียวดีเอ็นเอ กลไกการเลื่อนและการบิดเกลียวของเกลียว และการศึกษากำลังทางกายภาพที่เกี่ยวข้องกับกระบวนการเหล่านี้
ชีวฟิสิกส์ระดับโมเลกุลให้ความช่วยเหลืออย่างมากแก่อณูชีววิทยาในการศึกษาโครงสร้างของโมเลกุลโปรตีนด้วยความช่วยเหลือของการวิเคราะห์การเลี้ยวเบนของรังสีเอกซ์ ซึ่งใช้ครั้งแรกในปี 2473 โดย J. Bernal เป็นผลมาจากการใช้วิธีการทางกายภาพร่วมกับวิธีทางชีวเคมี (วิธีเอนไซม์) ที่เผยให้เห็นโครงสร้างโมเลกุลและลำดับของกรดอะมิโนในโปรตีนจำนวนหนึ่ง
การศึกษาด้วยกล้องจุลทรรศน์อิเล็กตรอนแบบสมัยใหม่ ซึ่งเผยให้เห็นการมีอยู่ของระบบเมมเบรนที่ซับซ้อนในเซลล์และออร์แกเนลล์ของมัน กระตุ้นความพยายามในการทำความเข้าใจโครงสร้างโมเลกุลของพวกมัน (ดูบทที่ 10 และ 11) มีการศึกษาองค์ประกอบทางเคมีของเมมเบรนและโดยเฉพาะอย่างยิ่งคุณสมบัติของไขมันในร่างกาย พบว่าหลังมีความสามารถในการออกซิเดชันมากเกินไปและปฏิกิริยาที่ไม่ใช่เอนไซม์ของการเกิดออกซิเดชันแบบลูกโซ่ (Yu. A. Vladimirov และ F. F. Litvin, 1959; B. N. Tarusov et al., 1960; I. I. Ivanov, 1967) ซึ่งนำไปสู่ความผิดปกติของเมมเบรน เพื่อศึกษาองค์ประกอบของเยื่อหุ้มเซลล์ก็เริ่มใช้วิธีการสร้างแบบจำลองทางคณิตศาสตร์ (V. Ts. Presman, 1964 - 1968; M. M. Shemyakin, 1967; Yu. A. Ovchinnikov, 1972)
ชีวฟิสิกส์ของเซลล์
เหตุการณ์สำคัญในประวัติศาสตร์ของชีวฟิสิกส์คือการก่อตัวของแนวคิดที่ชัดเจนเกี่ยวกับอุณหพลศาสตร์ของกระบวนการทางชีววิทยาในปี 1950 ซึ่งเป็นผลมาจากสมมติฐานเกี่ยวกับความเป็นไปได้ของการสร้างพลังงานอย่างอิสระในเซลล์ของสิ่งมีชีวิต ซึ่งขัดกับกฎข้อที่สองของอุณหพลศาสตร์ , หายไปในที่สุด การทำความเข้าใจการทำงานของกฎหมายนี้ในระบบทางชีววิทยานั้นเชื่อมโยงกับการแนะนำโดยนักวิทยาศาสตร์ชาวเบลเยียม I. Prigogine (1945) * ในอุณหพลศาสตร์ชีวภาพของแนวคิดของระบบเปิดที่แลกเปลี่ยนพลังงานและสสารกับสภาพแวดล้อมภายนอก Prigogine แสดงให้เห็นว่าเอนโทรปีบวกเกิดขึ้นในเซลล์ที่มีชีวิตในระหว่างกระบวนการทำงานตามกฎข้อที่สองของอุณหพลศาสตร์ สมการที่เขาแนะนำกำหนดเงื่อนไขภายใต้สภาวะที่เรียกว่าสถานะนิ่ง (ก่อนหน้านี้เรียกว่าสมดุลไดนามิก) ซึ่งปริมาณพลังงานอิสระ (negentropy) ที่เข้าสู่เซลล์ด้วยอาหารจะชดเชยการบริโภคและเอนโทรปีบวกคือ เอาท์พุท การค้นพบนี้ตอกย้ำแนวคิดทางชีววิทยาทั่วไปของการเชื่อมต่อที่แยกออกไม่ได้ระหว่างสภาพแวดล้อมภายนอกและภายในของเซลล์ เป็นจุดเริ่มต้นของการศึกษาอุณหพลศาสตร์ของระบบสิ่งมีชีวิตอย่างแท้จริง รวมถึงวิธีการสร้างแบบจำลอง (A. Burton, 1939; A. G. Pasynsky, 1967)
* (ทฤษฎีทั่วไปของระบบเปิดเสนอครั้งแรกโดย L. Bertalanffy ในปี 1932)
ตามหลักการพื้นฐานของไบโอเทอร์โมไดนามิกส์ เงื่อนไขที่จำเป็นสำหรับการดำรงอยู่ของชีวิตคือความไม่คงที่ในการพัฒนากระบวนการทางชีวเคมีของมัน สำหรับการดำเนินการซึ่งจำเป็นต้องประสานอัตราของปฏิกิริยาเมตาบอลิซึมจำนวนมาก บนพื้นฐานของเทอร์โมไดนามิกส์ทางชีวฟิสิกส์ใหม่นี้ มีแนวโน้มว่าจะแยกแยะปัจจัยภายนอกและภายในที่รับประกันการประสานกันของปฏิกิริยาและทำให้มีเสถียรภาพ ในช่วงสองทศวรรษที่ผ่านมา บทบาทสำคัญในการรักษาสภาวะนิ่งของระบบสารยับยั้งและโดยเฉพาะอย่างยิ่งสารต้านอนุมูลอิสระได้รับการเปิดเผย (B. N. Tarusov และ A. I. Zhuravlev, 1954, 1958) เป็นที่ยอมรับว่าความน่าเชื่อถือของการพัฒนาแบบอยู่กับที่นั้นสัมพันธ์กับปัจจัยด้านสิ่งแวดล้อม (อุณหภูมิ) และคุณสมบัติทางเคมีกายภาพของสภาพแวดล้อมของเซลล์
หลักการสมัยใหม่ของไบโอเทอร์โมไดนามิกส์ทำให้สามารถตีความกลไกการปรับตัวทางฟิสิกส์เคมีได้ จากข้อมูลของเรา การปรับตัวให้เข้ากับสภาพแวดล้อมสามารถเกิดขึ้นได้ก็ต่อเมื่อมีการเปลี่ยนแปลง ร่างกายสามารถสร้างความคงที่ในการพัฒนาปฏิกิริยาทางชีวเคมี (B.N. Tarusov, 1974) คำถามเกิดขึ้นจากการพัฒนาวิธีการใหม่ที่จะอนุญาตให้ประเมินสถานะนิ่งในร่างกายและทำนายการละเมิดที่เป็นไปได้ การนำหลักการทางไซเบอร์เนติกส์ของระบบการควบคุมตนเองมาใช้ในไบโอเทอร์โมไดนามิกส์ และการวิจัยเกี่ยวกับกระบวนการปรับตัวทางชีวภาพจะเป็นประโยชน์อย่างยิ่ง เป็นที่ชัดเจนว่าเพื่อแก้ปัญหาความเสถียรของสถานะนิ่งเป็นสิ่งสำคัญที่จะต้องคำนึงถึงปัจจัยที่เรียกว่าก่อกวนซึ่งรวมถึงโดยเฉพาะอย่างยิ่งปฏิกิริยาที่ไม่ใช่เอนไซม์ของออกซิเดชันของไขมัน เมื่อเร็ว ๆ นี้ การศึกษาเกี่ยวกับกระบวนการออกซิเดชันมากเกินไปในระยะไขมันของเซลล์ที่มีชีวิตและการเติบโตของผลิตภัณฑ์อนุมูลอิสระที่ละเมิดหน้าที่การกำกับดูแลของเยื่อหุ้มเซลล์ได้ขยายตัวขึ้น แหล่งที่มาของข้อมูลเกี่ยวกับกระบวนการเหล่านี้เป็นทั้งการตรวจหาอนุมูลเปอร์ออกไซด์และสารประกอบเปอร์ออกไซด์ของไบโอลิปิด (A. Tappel, 1965; I. I. Ivanov, 1965; E. B. Burlakova, 1967 และอื่นๆ) ในการตรวจหาอนุมูลอิสระ จะใช้ชีวเคมีลูมิเนสเซนซ์ ซึ่งเกิดขึ้นในไขมันของเซลล์ที่มีชีวิตในระหว่างการรวมตัวกันใหม่
บนพื้นฐานของความคิดทางเคมีกายภาพเกี่ยวกับความเสถียรของสภาวะคงตัว แนวคิดทางชีวฟิสิกส์เกิดขึ้นเกี่ยวกับการปรับตัวของพืชให้เข้ากับการเปลี่ยนแปลงในสภาพแวดล้อมที่เป็นการละเมิดระบบต้านอนุมูลอิสระที่ยับยั้ง (B. N. Tarusov, Ya. E. Doskoch, B. M. Kitlaev, A. M. Agaverdiev , 2511 - 2515) สิ่งนี้เปิดโอกาสในการประเมินคุณสมบัติต่างๆ เช่น การต้านทานความเย็นจัดและความทนทานต่อเกลือ ตลอดจนการทำนายที่เหมาะสมในการเลือกพืชผลทางการเกษตร
ในปี 1950 มีการค้นพบการเรืองแสงที่อ่อนแอเป็นพิเศษ - การเรืองแสงทางชีวภาพของวัตถุชีวภาพจำนวนหนึ่งในส่วนที่มองเห็นและอินฟราเรดของสเปกตรัม (B. N. Tarusov, A. I. Zhuravlev, A. I. Polivoda) สิ่งนี้เป็นไปได้อันเป็นผลมาจากการพัฒนาวิธีการลงทะเบียนฟลักซ์แสงที่อ่อนแอมากโดยใช้โฟโตมัลติเพลเยอร์ (L. A. Kubetsky, 1934) เป็นผลมาจากปฏิกิริยาทางชีวเคมีที่เกิดขึ้นในเซลล์ที่มีชีวิต ชีวเคมีลูมิเนสเซนซ์ทำให้สามารถตัดสินกระบวนการออกซิเดชันที่สำคัญในสายโซ่ถ่ายโอนอิเล็กตรอนระหว่างเอนไซม์ได้ การค้นพบและศึกษาชีวเคมีลูมิเนสเซนซ์มีความสำคัญมากทั้งทางทฤษฎีและทางปฏิบัติ ดังนั้น B. N. Tarusov และ Yu. B. Kudryashov ได้กล่าวถึงบทบาทที่ดีของผลิตภัณฑ์ของการเกิดออกซิเดชันของกรดไขมันไม่อิ่มตัวในกลไกของการเกิดสภาวะทางพยาธิสภาพที่พัฒนาภายใต้อิทธิพลของรังสีไอออไนซ์ในการก่อมะเร็งและการละเมิดอื่น ๆ ของการทำงานปกติ ของเซลล์
ในทศวรรษ 1950 ที่เกี่ยวข้องกับการพัฒนาอย่างรวดเร็วของฟิสิกส์นิวเคลียร์ รังสีชีววิทยา ซึ่งศึกษาผลกระทบทางชีวภาพของรังสีไอออไนซ์ ได้เกิดขึ้นจากชีวฟิสิกส์ การผลิตไอโซโทปกัมมันตภาพรังสี การสร้างอาวุธเทอร์โมนิวเคลียร์ เครื่องปฏิกรณ์ปรมาณู และการพัฒนารูปแบบอื่น ๆ ของการใช้พลังงานปรมาณูในทางปฏิบัติ ก่อให้เกิดปัญหาในการปกป้องสิ่งมีชีวิตจากอันตรายของรังสีไอออไนซ์ และพัฒนา พื้นฐานทางทฤษฎีในการป้องกันและรักษาโรคจากรังสี ในการทำเช่นนี้ ก่อนอื่นจำเป็นต้องค้นหาว่าส่วนประกอบใดของเซลล์และการเชื่อมโยงของการเผาผลาญอาหารที่เปราะบางที่สุด
วัตถุประสงค์ของการศึกษาชีวฟิสิกส์และรังสีชีววิทยาคือการอธิบายธรรมชาติของปฏิกิริยาเคมีเบื้องต้นที่เกิดขึ้นในพื้นผิวที่มีชีวิตภายใต้อิทธิพลของพลังงานรังสี ที่นี่เป็นสิ่งสำคัญที่ไม่เพียงแต่จะต้องเข้าใจกลไกของปรากฏการณ์นี้เท่านั้น แต่ยังต้องสามารถมีอิทธิพลต่อกระบวนการแลกเปลี่ยนพลังงานทางกายภาพเป็นพลังงานเคมี เพื่อลดสัมประสิทธิ์ของการกระทำที่ "มีประโยชน์" งานในทิศทางนี้เริ่มต้นโดยการศึกษาของโรงเรียน N. N. Semenov (1933) ในสหภาพโซเวียตและ D. Hinshelwood (1935) ในอังกฤษ
สถานที่สำคัญในการวิจัยทางรังสีวิทยาถูกครอบครองโดยการศึกษาระดับความต้านทานรังสีของสิ่งมีชีวิตต่างๆ พบว่าความต้านทานกัมมันตภาพรังสีที่เพิ่มขึ้น (เช่น ในสัตว์ฟันแทะในทะเลทราย) เกิดจากฤทธิ์ต้านอนุมูลอิสระสูงของไขมันในเยื่อหุ้มเซลล์ (M. Chang et al., 1964; N. K. Ogryzov et al., 1969) ปรากฎว่าโทโคฟีรอล วิตามินเค และสารประกอบไธโอมีบทบาทสำคัญในการสร้างคุณสมบัติต้านอนุมูลอิสระของระบบเหล่านี้ (II Ivanov et al., 1972) ในช่วงไม่กี่ปีที่ผ่านมา การศึกษากลไกการกลายพันธุ์ได้รับความสนใจอย่างมากเช่นกัน ด้วยเหตุนี้จึงทำการศึกษาผลของการแผ่รังสีไอออไนซ์ต่อพฤติกรรมของกรดนิวคลีอิกและโปรตีนในหลอดทดลอง เช่นเดียวกับในไวรัสและฟาจ (A. Gustafson, 1945 - 1950)
การต่อสู้เพื่อเพิ่มประสิทธิภาพในการป้องกันสารเคมี การค้นหาสารยับยั้งและหลักการในการยับยั้งที่มีประสิทธิภาพมากขึ้นยังคงเป็นภารกิจหลักของชีวฟิสิกส์ในทิศทางนี้
มีความคืบหน้าในการศึกษาสภาวะตื่นเต้นของพอลิเมอร์ชีวภาพ ซึ่งกำหนดกิจกรรมทางเคมีในระดับสูง ความสำเร็จมากที่สุดคือการศึกษาสภาวะตื่นเต้นที่เกิดขึ้นในขั้นตอนหลักของกระบวนการทางแสงชีวภาพ - การสังเคราะห์ด้วยแสงและการมองเห็น
ดังนั้นจึงมีส่วนสนับสนุนอย่างมากในการทำความเข้าใจการกระตุ้นเบื้องต้นของโมเลกุลของระบบเม็ดสีของพืช ความสำคัญอย่างยิ่งของการถ่ายโอน (การย้ายถิ่น) ของพลังงานของสภาวะที่ถูกกระตุ้นโดยไม่สูญเสียจากเม็ดสีที่ถูกกระตุ้นไปยังพื้นผิวอื่น ๆ ได้ถูกสร้างขึ้น มีบทบาทสำคัญในการพัฒนาแนวคิดเหล่านี้โดยงานเชิงทฤษฎีของ A. N. Terenin (1947 และต่อมา) A. A. Krasnovsky (1949) ค้นพบและศึกษาปฏิกิริยาของการลดโฟโตเคมีคอลแบบย้อนกลับของคลอโรฟิลล์และอะนาลอกของมัน ขณะนี้มีความเชื่อโดยทั่วไปว่าในอนาคตอันใกล้จะสามารถผลิตการสังเคราะห์ด้วยแสงภายใต้สภาวะที่ประดิษฐ์ขึ้นได้ (ดูบทที่ 5 ด้วย)
นักชีวฟิสิกส์ยังคงทำงานต่อไปเพื่อค้นหาธรรมชาติของการหดตัวของกล้ามเนื้อและกลไกของการกระตุ้นและการนำกระแสประสาท (ดูบทที่ 11) การวิจัยเกี่ยวกับกลไกของการเปลี่ยนจากสภาวะตื่นเต้นไปเป็นสภาวะปกติก็มีความสำคัญในปัจจุบันเช่นกัน สภาวะที่ตื่นเต้นได้รับการพิจารณาว่าเป็นผลจากปฏิกิริยาออโตคะตาไลติก และการยับยั้งถือเป็นผลสืบเนื่องมาจากการเคลื่อนตัวอย่างรวดเร็วของกิจกรรมการต้านอนุมูลอิสระที่ยับยั้งอันเป็นผลมาจากการจัดเรียงโมเลกุลใหม่ในสารประกอบ เช่น โทโคฟีรอล (I. I. Ivanov, O. R. Kols, 1966; O. R. คอลส์, 1970).
ปัญหาทั่วไปที่สำคัญที่สุดของชีวฟิสิกส์ยังคงเป็นความรู้เกี่ยวกับคุณสมบัติทางกายภาพและเคมีเชิงคุณภาพของสิ่งมีชีวิต คุณสมบัติต่างๆ เช่น ความสามารถของพอลิเมอร์ชีวภาพที่มีชีวิตในการผูกโพแทสเซียมหรือกระแสไฟฟ้าแบบโพลาไรซ์แบบเลือกสรรไม่สามารถรักษาไว้ได้ แม้จะกำจัดออกจากร่างกายอย่างระมัดระวังที่สุด ดังนั้นชีวฟิสิกส์ของเซลล์จึงยังคงพัฒนาเกณฑ์และวิธีการศึกษาสิ่งมีชีวิตอย่างต่อเนื่อง
แม้จะมีเยาวชนของชีววิทยาระดับโมเลกุล แต่ความคืบหน้าในด้านนี้เป็นสิ่งที่น่าทึ่งอย่างแท้จริง ในระยะเวลาอันสั้น ธรรมชาติของยีนและหลักการพื้นฐานของการจัดระเบียบ การสืบพันธุ์ และการทำงานได้ถูกสร้างขึ้น ยิ่งไปกว่านั้น ไม่เพียงแต่การสืบพันธุ์ของยีนในหลอดทดลองเท่านั้น แต่ยังได้ดำเนินการสังเคราะห์ยีนอย่างสมบูรณ์เป็นครั้งแรกด้วย รหัสพันธุกรรมได้รับการถอดรหัสอย่างสมบูรณ์ และปัญหาทางชีวภาพที่สำคัญที่สุดของความจำเพาะของการสังเคราะห์โปรตีนได้รับการแก้ไขแล้ว มีการระบุและศึกษาวิธีการและกลไกหลักของการสร้างโปรตีนในเซลล์ โครงสร้างหลักของ RNA การขนส่งจำนวนมาก ซึ่งเป็นโมเลกุลอะแดปเตอร์จำเพาะที่แปลภาษาของแม่แบบนิวคลีอิกเป็นภาษาของลำดับกรดอะมิโนของโปรตีนสังเคราะห์ ได้รับการพิจารณาอย่างสมบูรณ์แล้ว ลำดับกรดอะมิโนของโปรตีนหลายชนิดได้รับการถอดรหัสอย่างสมบูรณ์ และโครงสร้างเชิงพื้นที่ของโปรตีนบางชนิดได้รับการจัดตั้งขึ้น ทำให้สามารถอธิบายหลักการและรายละเอียดการทำงานของโมเลกุลของเอนไซม์ได้อย่างชัดเจน ทำการสังเคราะห์ทางเคมีของเอนไซม์ไรโบนิวคลีเอสตัวใดตัวหนึ่ง หลักการพื้นฐานของการจัดระเบียบของอนุภาคย่อยเซลล์ต่างๆ ไวรัสและฟาจจำนวนมากได้รับการจัดตั้งขึ้น และวิธีการหลักของการสร้างชีวภาพในเซลล์ได้รับการคลี่คลาย มีการค้นพบแนวทางในการทำความเข้าใจวิธีการควบคุมการทำงานของยีนและการอธิบายกลไกการกำกับดูแลของกิจกรรมที่สำคัญ แล้วรายการง่าย ๆ ของการค้นพบเหล่านี้บ่งชี้ว่าในช่วงครึ่งหลังของศตวรรษที่ 20 มีความก้าวหน้าอย่างมากในด้านชีววิทยา ซึ่งสาเหตุหลักมาจากการศึกษาเชิงลึกเกี่ยวกับโครงสร้างและหน้าที่ของโมเลกุลขนาดใหญ่ที่มีความสำคัญทางชีววิทยา - กรดนิวคลีอิกและโปรตีน
ความสำเร็จในอณูชีววิทยาได้ถูกนำมาใช้ในทางปฏิบัติในปัจจุบันและนำมาซึ่งผลลัพธ์ที่เป็นรูปธรรมในด้านการแพทย์ การเกษตร และบางอุตสาหกรรม ไม่ต้องสงสัยเลยว่าการกลับมาของวิทยาศาสตร์นี้จะเพิ่มขึ้นทุกวัน อย่างไรก็ตาม ผลลัพธ์หลักควรได้รับการพิจารณาว่าภายใต้อิทธิพลของความสำเร็จของอณูชีววิทยา ความมั่นใจในการดำรงอยู่ของความเป็นไปได้ที่ไม่ จำกัด ในการเผยความลับที่ลึกลับที่สุดของชีวิตได้แข็งแกร่งขึ้น
เห็นได้ชัดว่าในอนาคตจะมีการเปิดวิธีการใหม่ในการศึกษารูปแบบทางชีวภาพของการเคลื่อนที่ของสสาร - ชีววิทยาจะย้ายจากระดับโมเลกุลไปสู่ระดับอะตอม อย่างไรก็ตาม ในตอนนี้ อาจไม่มีนักวิจัยแม้แต่คนเดียวที่สามารถทำนายพัฒนาการทางอณูชีววิทยาได้อย่างสมจริง แม้กระทั่งในอีก 20 ปีข้างหน้า
ความก้าวหน้าในการศึกษากรดนิวคลีอิกและการสังเคราะห์โปรตีนได้นำไปสู่การพัฒนาวิธีการที่มีความสำคัญในทางปฏิบัติอย่างมากในด้านการแพทย์ การเกษตร และอุตสาหกรรมอื่นๆ จำนวนหนึ่ง
หลังจากศึกษารหัสพันธุกรรมและหลักการพื้นฐานของการจัดเก็บและใช้ข้อมูลทางพันธุกรรมแล้ว การพัฒนาของอณูชีววิทยาก็หยุดนิ่ง เนื่องจากไม่มีวิธีการใดที่ทำให้สามารถควบคุมยีน แยกออก และเปลี่ยนแปลงยีนได้ การเกิดขึ้นของวิธีการเหล่านี้เกิดขึ้นในปี 1970-1980 สิ่งนี้เป็นแรงผลักดันอันทรงพลังในการพัฒนาสาขาวิทยาศาสตร์นี้ ซึ่งยังคงเฟื่องฟูมาจนถึงทุกวันนี้ อย่างแรกเลย วิธีการเหล่านี้เกี่ยวข้องกับการได้มาซึ่งยีนแต่ละตัวและการนำพวกมันเข้าสู่เซลล์ของสิ่งมีชีวิตอื่น (การโคลนโมเลกุลและการถ่ายยีน, PCR) รวมถึงวิธีการกำหนดลำดับนิวคลีโอไทด์ในยีน (การจัดลำดับ DNA และ RNA) วิธีการเหล่านี้จะกล่าวถึงในรายละเอียดเพิ่มเติมด้านล่าง เราจะเริ่มด้วยวิธีพื้นฐานที่ง่ายที่สุด นั่นคือ อิเล็กโตรโฟรีซิส จากนั้นไปยังวิธีที่ซับซ้อนมากขึ้น
ดีเอ็นเออิเล็กโทรโฟเรซิส
เป็นวิธีการพื้นฐานในการทำงานกับ DNA ซึ่งใช้ร่วมกับวิธีอื่นๆ เกือบทั้งหมดเพื่อแยกโมเลกุลที่ต้องการและวิเคราะห์ผลลัพธ์ เจลอิเล็กโตรโฟรีซิสใช้เพื่อแยกชิ้นส่วนดีเอ็นเอตามความยาว DNA เป็นกรด โมเลกุลของมันมีกรดฟอสฟอริกตกค้าง ซึ่งแยกโปรตอนออกและได้รับประจุลบ (รูปที่ 1)
ดังนั้น ในสนามไฟฟ้า โมเลกุลดีเอ็นเอจะเคลื่อนเข้าหาขั้วบวก ซึ่งเป็นอิเล็กโทรดที่มีประจุบวก สิ่งนี้เกิดขึ้นในสารละลายอิเล็กโทรไลต์ที่มีไอออนพาหะประจุ เนื่องจากสารละลายนี้นำกระแส ในการแยกชิ้นส่วน ใช้เจลหนาแน่นที่ทำจากโพลีเมอร์ (agarose หรือ polyacrylamide) โมเลกุลของดีเอ็นเอ "พัวพัน" เข้าไปยิ่งนาน ดังนั้นโมเลกุลที่ยาวที่สุดจะเคลื่อนที่ช้าที่สุดและสั้นที่สุด - เร็วที่สุด (รูปที่ 2) ก่อนหรือหลังอิเล็กโตรโฟรีซิส เจลจะได้รับการบำบัดด้วยสีย้อมที่ผูกมัดกับ DNA และเรืองแสงในแสงอัลตราไวโอเลต และได้รูปแบบของแถบในเจล (ดูรูปที่ 3) ในการกำหนดความยาวของชิ้นส่วนดีเอ็นเอในตัวอย่าง จะถูกนำไปเปรียบเทียบกับเครื่องหมาย กล่าวคือ ชุดของชิ้นส่วนที่มีความยาวมาตรฐานวางขนานกันบนเจลเดียวกัน (รูปที่ 4)



เครื่องมือที่สำคัญที่สุดในการทำงานกับ DNA คือเอนไซม์ที่ทำการเปลี่ยนแปลงของ DNA ในเซลล์ของสิ่งมีชีวิต: DNA polymerase, DNA ligases, และจำกัดเอนโดนิวคลีเอสหรือเอนไซม์จำกัด ดีเอ็นเอโพลีเมอเรสมีการสังเคราะห์แม่แบบ DNA ซึ่งช่วยให้สามารถขยายพันธุ์ DNA ในหลอดทดลองได้ ดีเอ็นเอ ไลกาซีสเย็บโมเลกุล DNA เข้าด้วยกันหรือรักษาช่องว่างในพวกมัน ข้อ จำกัด เอ็นโดนิวคลีเอส, หรือ ข้อจำกัดตัดโมเลกุล DNA ตามลำดับที่กำหนดไว้อย่างเคร่งครัด ซึ่งช่วยให้คุณตัดชิ้นส่วนแต่ละส่วนออกจากมวลรวมของ DNA ได้ ชิ้นส่วนเหล่านี้ในบางกรณีอาจมียีนแต่ละตัว
ข้อจำกัด
ลำดับที่รับรู้โดยเอ็นไซม์การจำกัดนั้นมีความสมมาตร และการแตกหักอาจเกิดขึ้นได้ในช่วงกลางของลำดับดังกล่าวหรือมีการเปลี่ยนแปลง (ในที่เดียวกันในสายดีเอ็นเอทั้งสองสาย) โครงร่างการดำเนินการของข้อ จำกัด ประเภทต่าง ๆ แสดงในรูปที่ 1. ในกรณีแรกจะได้สิ่งที่เรียกว่า "ทื่อ" และในกรณีที่สอง - ปลาย "เหนียว" ในกรณีของปลาย "เหนียว" ของด้านล่าง ห่วงโซ่จะสั้นกว่าส่วนอื่น ๆ ส่วนที่เป็นเกลียวเดี่ยวจะถูกสร้างขึ้นด้วยลำดับสมมาตรที่เหมือนกันที่ปลายทั้งสองแบบ
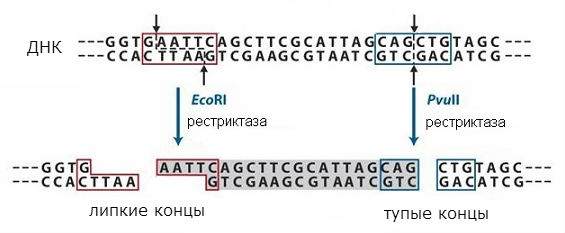
ลำดับสุดท้ายจะเหมือนกันเมื่อ DNA ใดๆ ถูกตัดแยกด้วยเอ็นไซม์การจำกัดที่กำหนดและสามารถเข้าร่วมใหม่ได้เนื่องจากมีลำดับที่เสริมกัน พวกเขาสามารถผูกกับ DNA ligase เพื่อสร้างโมเลกุลเดี่ยว ดังนั้นจึงเป็นไปได้ที่จะรวมชิ้นส่วนของ DNA ที่แตกต่างกันสองชิ้นเข้าด้วยกันและได้สิ่งที่เรียกว่า ดีเอ็นเอลูกผสม. วิธีนี้ใช้ในวิธีการโคลนโมเลกุล ซึ่งทำให้สามารถรับยีนแต่ละตัวและนำเข้าสู่เซลล์ที่สามารถสร้างโปรตีนที่เข้ารหัสในยีนได้
การโคลนโมเลกุล
การโคลนโมเลกุลใช้โมเลกุลดีเอ็นเอสองโมเลกุล - เม็ดมีดที่มียีนที่น่าสนใจ และ เวกเตอร์- DNA ทำหน้าที่เป็นพาหะ เม็ดมีดถูก "เย็บ" เข้าไปในเวกเตอร์ด้วยความช่วยเหลือของเอ็นไซม์ ได้โมเลกุลดีเอ็นเอลูกผสมใหม่ จากนั้นโมเลกุลนี้จะถูกนำเข้าสู่เซลล์เจ้าบ้าน และเซลล์เหล่านี้ก่อตัวเป็นโคโลนีบนอาหารเลี้ยงเชื้อ อาณานิคมเป็นลูกหลานของเซลล์เดียว กล่าวคือ โคลน เซลล์ทั้งหมดของอาณานิคมมีลักษณะทางพันธุกรรมเหมือนกันและมี DNA ลูกผสมเดียวกัน ดังนั้นคำว่า "การโคลนโมเลกุล" นั่นคือการได้รับโคลนของเซลล์ที่มีชิ้นส่วนดีเอ็นเอที่เราสนใจ หลังจากที่ได้โคโลนีที่มีส่วนแทรกที่เราสนใจแล้ว ก็เป็นไปได้ที่จะระบุลักษณะเฉพาะส่วนแทรกนี้ด้วยวิธีการต่างๆ เช่น เพื่อกำหนดลำดับที่แน่นอนของเม็ดมีด เซลล์ยังสามารถผลิตโปรตีนที่เข้ารหัสโดยส่วนแทรกได้หากมียีนที่ใช้งานได้
เมื่อมีการนำโมเลกุลลูกผสมเข้าไปในเซลล์ การเปลี่ยนแปลงทางพันธุกรรมของเซลล์เหล่านี้จะเกิดขึ้น การแปลงร่าง- กระบวนการดูดซึมโดยเซลล์ของสิ่งมีชีวิตของโมเลกุล DNA อิสระจากสิ่งแวดล้อมและการรวมเข้ากับจีโนมซึ่งนำไปสู่การปรากฏตัวในเซลล์ของลักษณะที่สืบทอดใหม่ของมันซึ่งเป็นลักษณะของผู้บริจาคสิ่งมีชีวิตของ DNA . ตัวอย่างเช่น หากโมเลกุลที่แทรกมียีนที่ดื้อต่อยาปฏิชีวนะแอมพิซิลลิน แบคทีเรียที่เปลี่ยนแปลงแล้วจะเติบโตต่อหน้า ก่อนการเปลี่ยนแปลงแอมพิซิลลินทำให้เกิดความตายนั่นคือสัญญาณใหม่ปรากฏขึ้นในเซลล์ที่เปลี่ยนรูป
เวกเตอร์
เวกเตอร์ต้องมีคุณสมบัติหลายประการ:
ประการแรก มันเป็นโมเลกุล DNA ที่ค่อนข้างเล็กที่จะถูกจัดการอย่างง่ายดาย
ประการที่สอง เพื่อให้ DNA ถูกรักษาและทำซ้ำในเซลล์ จะต้องมีลำดับที่แน่นอนเพื่อให้แน่ใจว่าการจำลองแบบของมัน (ที่มาของการจำลองหรือที่มาของการจำลองแบบ)
ประการที่สาม จะต้องมี ยีนเครื่องหมายซึ่งทำให้แน่ใจถึงการเลือกเฉพาะเซลล์ที่เวกเตอร์เข้าไป โดยปกติสิ่งเหล่านี้คือยีนต้านทานยาปฏิชีวนะ - จากนั้นเมื่อมียาปฏิชีวนะ เซลล์ทั้งหมดที่ไม่มีพาหะนำโรคจะตาย
การโคลนยีนส่วนใหญ่มักเกิดขึ้นในเซลล์แบคทีเรีย เนื่องจากง่ายต่อการปลูกฝังและเพิ่มจำนวนอย่างรวดเร็ว ในเซลล์แบคทีเรีย มักจะมีโมเลกุลดีเอ็นเอทรงกลมขนาดใหญ่หนึ่งโมเลกุล ซึ่งมีความยาวหลายล้านคู่เบส ซึ่งบรรจุยีนทั้งหมดที่จำเป็นสำหรับแบคทีเรีย นั่นคือโครโมโซมของแบคทีเรีย นอกจากนี้ในแบคทีเรียบางชนิดยังมี DNA ทรงกลมขนาดเล็ก (หลายพันคู่เบส) เรียกว่า พลาสมิด(รูปที่ 2). เช่นเดียวกับ DNA หลักที่มีลำดับนิวคลีโอไทด์ที่ให้ความสามารถของ DNA ในการทำซ้ำ (ori) พลาสมิดทำซ้ำโดยไม่ขึ้นกับ DNA หลัก (โครโมโซม) ดังนั้นจึงมีอยู่ในเซลล์ในสำเนาจำนวนมาก พลาสมิดเหล่านี้จำนวนมากมียีนต้านทานยาปฏิชีวนะ ซึ่งทำให้สามารถแยกแยะเซลล์ที่มีพลาสมิดออกจากเซลล์ปกติได้ โดยทั่วไป พลาสมิดที่มียีนสองตัวที่ให้การดื้อต่อยาปฏิชีวนะ 2 ชนิด เช่น เตตราไซคลินและอะไมซิลิน มีวิธีการง่ายๆ ในการแยกพลาสมิด DNA ออกจาก DNA ของโครโมโซมหลักของแบคทีเรีย

ความสำคัญของการถ่ายทอดทางพันธุกรรม
การถ่ายทอดยีนจากสิ่งมีชีวิตหนึ่งไปยังอีกสิ่งมีชีวิตหนึ่งเรียกว่า การดัดแปลงพันธุกรรมและสิ่งมีชีวิตดัดแปลงดังกล่าว - ดัดแปลงพันธุกรรม. วิธีการถ่ายโอนยีนไปยังเซลล์จุลินทรีย์นั้นใช้เพื่อเตรียมโปรตีนรีคอมบิแนนท์สำหรับยา โดยเฉพาะอย่างยิ่ง โปรตีนของมนุษย์ที่ไม่ก่อให้เกิดการปฏิเสธภูมิคุ้มกัน - อินเตอร์เฟอรอน อินซูลิน และฮอร์โมนโปรตีนอื่นๆ ปัจจัยการเจริญเติบโตของเซลล์ เช่นเดียวกับโปรตีนสำหรับการผลิต วัคซีน. ในกรณีที่ซับซ้อนมากขึ้น เมื่อการดัดแปลงโปรตีนดำเนินการอย่างถูกต้องเฉพาะในเซลล์ยูคาริโอต การเพาะเลี้ยงเซลล์ดัดแปลงพันธุกรรมหรือสัตว์ดัดแปลงพันธุกรรมถูกนำมาใช้ โดยเฉพาะอย่างยิ่ง ปศุสัตว์ (ส่วนใหญ่เป็นแพะ) ซึ่งหลั่งโปรตีนที่จำเป็นลงในนม หรือแยกโปรตีนออกจากเลือดของพวกมัน นี่คือวิธีการรับแอนติบอดี ปัจจัยการแข็งตัวของเลือด และโปรตีนอื่นๆ วิธีการแปลงพันธุ์ผลิตพืชที่เพาะปลูกที่ทนทานต่อสารกำจัดวัชพืชและแมลงศัตรูพืช และมีคุณสมบัติที่มีประโยชน์อื่นๆ การใช้จุลินทรีย์แปลงพันธุ์เพื่อทำให้น้ำเสียบริสุทธิ์และต่อสู้กับมลภาวะ มีแม้กระทั่งจุลินทรีย์แปลงพันธุ์ที่สามารถย่อยสลายน้ำมันได้ นอกจากนี้ เทคโนโลยีดัดแปรพันธุกรรมเป็นสิ่งที่ขาดไม่ได้ในการวิจัยทางวิทยาศาสตร์ - การพัฒนาทางชีววิทยาในปัจจุบันเป็นสิ่งที่คิดไม่ถึงหากไม่มีการใช้การดัดแปลงและถ่ายทอดยีนเป็นประจำ
เทคโนโลยีการโคลนโมเลกุล
เม็ดมีด
เพื่อให้ได้ยีนแต่ละตัวจากสิ่งมีชีวิตใดๆ DNA โครโมโซมทั้งหมดจะถูกแยกออกจากยีนนั้นและแยกออกด้วยเอ็นไซม์จำกัดหนึ่งหรือสองตัว เอ็นไซม์ได้รับการคัดเลือกเพื่อไม่ให้ตัดยีนที่เราสนใจ แต่จะแตกไปตามขอบของมัน และในพลาสมิด DNA จะทำให้ยีนต้านทานตัวใดตัวหนึ่งแตก เช่น แอมพิซิลลิน
กระบวนการโคลนโมเลกุลประกอบด้วยขั้นตอนต่อไปนี้:
ตัดและเย็บ - การสร้างโมเลกุลลูกผสมเดี่ยวจากเม็ดมีดและเวกเตอร์
การเปลี่ยนแปลงคือการนำโมเลกุลลูกผสมเข้าสู่เซลล์
การเลือก - การเลือกเซลล์ที่ได้รับเวกเตอร์พร้อมส่วนแทรก
ตัดและเย็บ
พลาสมิดดีเอ็นเอได้รับการบำบัดด้วยเอ็นไซม์การจำกัดแบบเดียวกัน และจะกลายเป็นโมเลกุลเชิงเส้นถ้าเลือกเอ็นไซม์การจำกัดดังกล่าวซึ่งทำให้เกิดการแตกตัวในพลาสมิด 1 ครั้ง เป็นผลให้ปลายเหนียวเดียวกันปรากฏขึ้นที่ส่วนปลายของชิ้นส่วนดีเอ็นเอที่เกิดขึ้นทั้งหมด เมื่ออุณหภูมิลดลง ปลายเหล่านี้จะเชื่อมติดกันแบบสุ่มและผูกมัดด้วย DNA ligase (ดูรูปที่ 3)

ได้ส่วนผสมของ DNA ทรงกลมที่มีองค์ประกอบต่างกัน: บางส่วนจะมีลำดับ DNA บางอย่างของ DNA โครโมโซมที่เชื่อมต่อกับ DNA ของแบคทีเรีย ส่วนอื่นๆ จะมีชิ้นส่วนของ DNA ของโครโมโซมที่เชื่อมต่อเข้าด้วยกัน และส่วนอื่นๆ จะมีพลาสมิดแบบรีดิวซ์หรือไดเมอร์ (รูปที่ 4).

การเปลี่ยนแปลง
ถัดไป ผสมนี้จะดำเนินการ การเปลี่ยนแปลงทางพันธุกรรมแบคทีเรียที่ไม่มีพลาสมิด การแปลงร่าง- กระบวนการดูดซึมโดยเซลล์ของสิ่งมีชีวิตของโมเลกุล DNA อิสระจากสิ่งแวดล้อมและการรวมเข้ากับจีโนมซึ่งนำไปสู่การปรากฏตัวในเซลล์ของลักษณะที่สืบทอดใหม่ของมันซึ่งเป็นลักษณะของผู้บริจาคสิ่งมีชีวิตของ DNA . พลาสมิดเพียงหนึ่งตัวเท่านั้นที่สามารถป้อนและเพิ่มจำนวนในแต่ละเซลล์ เซลล์ดังกล่าววางอยู่บนอาหารที่มีสารอาหารที่เป็นของแข็งซึ่งมียาปฏิชีวนะเตตราไซคลิน เซลล์ที่ไม่ได้รับพลาสมิดจะไม่เติบโตบนตัวกลางนี้ และเซลล์ที่มีโคโลนีจากรูปแบบพลาสมิด ซึ่งแต่ละเซลล์มีลูกหลานของเซลล์เพียงเซลล์เดียว กล่าวคือ เซลล์ทั้งหมดในอาณานิคมมีพลาสมิดตัวเดียวกัน (ดูรูปที่ 5)

การคัดเลือก
ขั้นต่อไป ภารกิจคือการแยกเฉพาะเซลล์ที่เวกเตอร์ที่มีการแทรกเข้าไป และเพื่อแยกความแตกต่างออกจากเซลล์ที่มีเฉพาะเวกเตอร์ที่ไม่มีส่วนแทรก หรือไม่ถือเวกเตอร์เลย ขั้นตอนการเลือกเซลล์ที่ถูกต้องนี้เรียกว่า การเลือก. สำหรับสิ่งนี้ สมัคร เครื่องหมายเลือก- โดยปกติยีนดื้อยาปฏิชีวนะในเวกเตอร์และ สื่อคัดสรรที่มียาปฏิชีวนะหรือสารเฉพาะอื่นๆ
ในตัวอย่างที่เรากำลังพิจารณา เซลล์จากอาณานิคมที่ปลูกต่อหน้าแอมพิซิลลินนั้นได้รับการเพาะเลี้ยงย่อยบนสื่อสองชนิด: เซลล์แรกประกอบด้วยแอมพิซิลลิน และเซลล์ที่สองประกอบด้วยเตตราไซคลิน โคโลนีที่มีเพียงพลาสมิดจะเติบโตบนตัวกลางทั้งสอง ในขณะที่โคโลนีที่มี DNA โครโมโซมที่แทรกอยู่ในพลาสมิดจะไม่เติบโตบนอาหารที่มีเตตราไซคลิน (รูปที่ 5) ในหมู่พวกเขา ยีนที่เราสนใจจะถูกเลือกโดยวิธีพิเศษ ปลูกในปริมาณที่เพียงพอ และแยก DNA พลาสมิดออก จากนั้น ใช้ข้อจำกัดเดียวกันกับที่ใช้ในการรับ DNA รีคอมบิแนนท์ ยีนแต่ละตัวที่สนใจจะถูกตัดออก ดีเอ็นเอของยีนนี้สามารถใช้เพื่อกำหนดลำดับของนิวคลีโอไทด์ นำเข้าสู่สิ่งมีชีวิตใดๆ เพื่อให้ได้คุณสมบัติใหม่หรือสังเคราะห์โปรตีนที่ต้องการ วิธีการแยกยีนนี้เรียกว่า การโคลนโมเลกุล.
โปรตีนเรืองแสง
สะดวกในการใช้โปรตีนเรืองแสงเป็นยีนเครื่องหมายในการศึกษาสิ่งมีชีวิตที่มียูคาริโอต ยีนของโปรตีนเรืองแสงตัวแรก โปรตีนเรืองแสงสีเขียว (GFP)ถูกแยกออกจากแมงกะพรุน Aqeuorea victoria และนำเข้าสู่สิ่งมีชีวิตจำลองต่างๆ (ดูรูปที่ 6) ในปี 2008 O. Shimomura, M. Chalfi และ R. Tsien ได้รับรางวัลโนเบลสำหรับการค้นพบและการประยุกต์ใช้โปรตีนนี้

จากนั้นจึงแยกยีนสำหรับโปรตีนเรืองแสงอื่นๆ - แดง น้ำเงิน เหลือง ยีนเหล่านี้ได้รับการดัดแปลงเทียมเพื่อผลิตโปรตีนที่มีคุณสมบัติตามที่ต้องการ ความหลากหลายของโปรตีนเรืองแสงแสดงในรูปที่ 7 ซึ่งแสดงจานเพาะเชื้อที่มีแบคทีเรียที่มียีนสำหรับโปรตีนเรืองแสงต่างๆ

การใช้โปรตีนเรืองแสง
ยีนโปรตีนเรืองแสงสามารถหลอมรวมกับยีนของโปรตีนอื่นๆ ได้ จากนั้นในระหว่างการแปล โปรตีนเดี่ยวจะถูกสร้างขึ้น - โปรตีนหลอมรวมการแปลหรือ ฟิวชั่น(ฟิวชันโปรตีน) ซึ่งเรืองแสง ดังนั้นจึงเป็นไปได้ที่จะศึกษาตัวอย่างเช่นการแปล (ตำแหน่ง) ของโปรตีนใด ๆ ที่น่าสนใจในเซลล์การเคลื่อนไหวของพวกมัน การใช้การแสดงออกของโปรตีนเรืองแสงเฉพาะในเซลล์บางประเภทเท่านั้นจึงเป็นไปได้ที่จะทำเครื่องหมายเซลล์ประเภทนี้ในสิ่งมีชีวิตหลายเซลล์ (ดูรูปที่ 8 - สมองของหนูซึ่งเซลล์ประสาทแต่ละเซลล์มีสีต่างกันเนื่องจากการรวมกันของโปรตีนเรืองแสง ยีน) โปรตีนเรืองแสงเป็นเครื่องมือที่จำเป็นในอณูชีววิทยาสมัยใหม่

PCR
อีกวิธีหนึ่งในการได้มาซึ่งยีนเรียกว่า ปฏิกิริยาลูกโซ่โพลีเมอเรส (PCR). มันขึ้นอยู่กับความสามารถของ DNA polymerase ในการทำให้สายที่สองของ DNA สมบูรณ์ตามสายคู่สม ดังที่เกิดขึ้นในเซลล์ระหว่างการจำลองแบบของ DNA
ต้นกำเนิดของการจำลองแบบในวิธีนี้ได้มาจาก DNA เล็กๆ สองชิ้นที่เรียกว่า เมล็ดพืชหรือ ไพรเมอร์. ไพรเมอร์เหล่านี้เป็นส่วนเติมเต็มของยีนที่สนใจบน DNA สองสาย อย่างแรก โครโมโซม DNA ที่จะแยกยีนออกจากกันนั้นผสมกับเมล็ดพืชและให้ความร้อนถึง 99 o C ซึ่งนำไปสู่การแตกของพันธะไฮโดรเจนและความแตกต่างของสาย DNA หลังจากนั้นอุณหภูมิจะลดลงเหลือ 50-70 ประมาณ C (ขึ้นอยู่กับความยาวและลำดับของเมล็ด) ภายใต้เงื่อนไขเหล่านี้ ไพรเมอร์จะถูกยึดติดกับบริเวณเสริมของ DNA โครโมโซม ทำให้เกิดเกลียวคู่ปกติ (ดูรูปที่ 9) หลังจากนั้นจะมีการเพิ่มส่วนผสมของนิวคลีโอไทด์ทั้งสี่ที่จำเป็นสำหรับการสังเคราะห์ DNA และ DNA polymerase เอ็นไซม์ยืดไพรเมอร์โดยการสร้าง DNA ที่มีเกลียวสองเส้นจากจุดที่เกาะติดกับไพรเมอร์ กล่าวคือ ตั้งแต่ปลายยีนจนถึงปลายโมเลกุลโครโมโซมสายเดี่ยว 
หากส่วนผสมถูกทำให้ร้อนอีกครั้ง โครโมโซมและสายที่สังเคราะห์ขึ้นใหม่จะกระจายตัว หลังจากเย็นตัวลงเมล็ดจะเข้าร่วมอีกครั้งซึ่งเกินขนาดมาก (ดูรูปที่ 10)

บนสายโซ่ที่สังเคราะห์ขึ้นใหม่ พวกมันจะไม่เชื่อมติดกันจนถึงจุดสิ้นสุดที่การสังเคราะห์ครั้งแรกเริ่มต้น แต่กับสายตรงข้าม เนื่องจากสายโซ่ DNA นั้นตรงกันข้ามกัน ดังนั้น ในรอบที่สองของการสังเคราะห์ เฉพาะลำดับที่สอดคล้องกับยีนเท่านั้นที่จะเสร็จสมบูรณ์บนสายโซ่ดังกล่าว (ดูรูปที่ 11)

วิธีนี้ใช้ DNA polymerase จากแบคทีเรีย thermophilic ที่สามารถทนต่อการเดือดและทำงานที่อุณหภูมิ 70-80 ° C ได้ ไม่จำเป็นต้องเติมทุกครั้ง แต่พอเพิ่มเข้าไปตอนเริ่มการทดลอง โดยการทำซ้ำขั้นตอนการให้ความร้อนและความเย็นในลำดับเดียวกัน เราสามารถเพิ่มจำนวนลำดับในแต่ละรอบได้เป็นสองเท่า โดยจะล้อมรอบปลายทั้งสองข้างด้วยเมล็ดที่นำเข้า (ดูรูปที่ 12)

หลังจากประมาณ 25 รอบดังกล่าว จำนวนสำเนาของยีนจะเพิ่มขึ้นมากกว่าหนึ่งล้านเท่า ปริมาณดังกล่าวสามารถแยกออกจาก DNA โครโมโซมที่ใส่เข้าไปในหลอดทดลองได้อย่างง่ายดายและใช้เพื่อวัตถุประสงค์ต่างๆ
ลำดับดีเอ็นเอ
ความสำเร็จที่สำคัญอีกประการหนึ่งคือการพัฒนาวิธีการกำหนดลำดับของนิวคลีโอไทด์ใน DNA - ลำดับดีเอ็นเอ(จากลำดับภาษาอังกฤษ - ลำดับ). ในการทำเช่นนี้ จำเป็นต้องได้รับยีนที่บริสุทธิ์จาก DNA อื่นโดยใช้วิธีใดวิธีหนึ่งที่อธิบายไว้ จากนั้นสายดีเอ็นเอจะถูกแยกออกโดยการให้ความร้อนและไพรเมอร์ที่ติดฉลากด้วยฟอสฟอรัสกัมมันตภาพรังสีหรือฉลากเรืองแสงจะถูกเพิ่มเข้าไป โปรดทราบว่ามีการใช้เมล็ดพันธุ์หนึ่งเมล็ด ประกอบกับห่วงโซ่เดียว จากนั้นเติม DNA polymerase และส่วนผสมของ 4 นิวคลีโอไทด์ ส่วนผสมดังกล่าวแบ่งออกเป็น 4 ส่วนและนิวคลีโอไทด์หนึ่งตัวถูกเติมในแต่ละส่วน ดัดแปลงเพื่อไม่ให้มีหมู่ไฮดรอกซิลในอะตอมที่สามของดีออกซีไรโบส หากนิวคลีโอไทด์ดังกล่าวรวมอยู่ในสายดีเอ็นเอสังเคราะห์ การยืดตัวของนิวคลีโอไทด์จะไม่สามารถดำเนินต่อไปได้เพราะ พอลิเมอเรสจะไม่มีส่วนที่จะเกาะนิวคลีโอไทด์ต่อไป ดังนั้นการสังเคราะห์ดีเอ็นเอหลังจากการรวมนิวคลีโอไทด์ดังกล่าวจึงถูกขัดจังหวะ นิวคลีโอไทด์เหล่านี้เรียกว่าไดดีออกซีนิวคลีโอไทด์ถูกเติมเข้าไปน้อยกว่าปกติมาก ดังนั้นการยุติสายโซ่เกิดขึ้นเป็นครั้งคราวเท่านั้นและในแต่ละสายในที่ต่างกัน ผลที่ได้คือส่วนผสมของสายโซ่ที่มีความยาวต่างกัน โดยแต่ละสายมีนิวคลีโอไทด์เหมือนกันที่ปลาย ดังนั้น ความยาวสายโซ่จึงสอดคล้องกับหมายเลขนิวคลีโอไทด์ในลำดับที่ทำการศึกษา ตัวอย่างเช่น หากเรามีแอดีนิล ไดดีออกซีนิวคลีโอไทด์ และสายที่เป็นผลลัพธ์จะมีความยาวนิวคลีโอไทด์ 2, 7 และ 12 ตัว จากนั้นอะดีนีนจะอยู่ในตำแหน่งที่สอง เจ็ด และสิบสอง ในยีน ส่วนผสมที่เป็นผลลัพธ์ของสายโซ่สามารถแยกออกได้ง่ายตามขนาดโดยใช้อิเล็กโตรโฟรีซิส และสามารถระบุสายที่สังเคราะห์ได้โดยใช้กัมมันตภาพรังสีบนฟิล์มเอ็กซ์เรย์ (ดูรูปที่ 10)

ปรากฎภาพที่ด้านล่างของภาพที่เรียกว่า radioautograph เมื่อเลื่อนจากล่างขึ้นบนและอ่านตัวอักษรเหนือคอลัมน์ของแต่ละโซน เราจะได้ลำดับนิวคลีโอไทด์ที่แสดงในรูปทางด้านขวาของลายเซ็น ปรากฎว่าการสังเคราะห์หยุดไม่เพียงแค่โดยไดดีออกซีนิวคลีโอไทด์เท่านั้น แต่ยังรวมถึงนิวคลีโอไทด์ด้วยซึ่งกลุ่มเคมีบางกลุ่ม เช่น สีย้อมเรืองแสง ติดอยู่ในตำแหน่งที่สามของน้ำตาล หากนิวคลีโอไทด์แต่ละอันติดฉลากด้วยสีย้อมของตัวเอง โซนที่ได้จากการแยกโซ่ที่สังเคราะห์แล้วจะเรืองแสงด้วยแสงที่แตกต่างกัน สิ่งนี้ทำให้สามารถทำปฏิกิริยาในหลอดทดลองหนึ่งหลอดพร้อมกันสำหรับนิวคลีโอไทด์ทั้งหมด และโดยการแยกสายที่เป็นผลลัพธ์ตามความยาว เพื่อระบุนิวคลีโอไทด์ตามสี (ดูรูปที่ 11)

วิธีการดังกล่าวทำให้สามารถกำหนดลำดับไม่เฉพาะของยีนแต่ละตัวเท่านั้น แต่ยังอ่านจีโนมทั้งหมดได้ด้วย ปัจจุบันได้มีการพัฒนาวิธีการที่รวดเร็วยิ่งขึ้นในการกำหนดลำดับนิวคลีโอไทด์ในยีน หากจีโนมมนุษย์กลุ่มแรกถูกถอดรหัสโดยสมาคมระหว่างประเทศขนาดใหญ่โดยใช้วิธีที่กำหนดครั้งแรกใน 12 ปี ครั้งที่สอง โดยใช้วิธีที่สอง ใน 3 ปี ตอนนี้สามารถทำได้ในหนึ่งเดือน วิธีนี้ช่วยให้คุณทำนายความโน้มเอียงของบุคคลต่อโรคต่างๆ และใช้มาตรการล่วงหน้าเพื่อหลีกเลี่ยง
31.2
เพื่อเพื่อน!
อ้างอิง
อณูชีววิทยาเติบโตจากชีวเคมีในเดือนเมษายน พ.ศ. 2496 ลักษณะที่ปรากฏเกี่ยวข้องกับชื่อของ James Watson และ Francis Crick ผู้ค้นพบโครงสร้างของโมเลกุลดีเอ็นเอ การค้นพบนี้เกิดขึ้นได้จากการศึกษาพันธุศาสตร์ แบคทีเรีย และชีวเคมีของไวรัส อาชีพนักชีววิทยาระดับโมเลกุลยังไม่แพร่หลายนัก แต่ปัจจุบันบทบาทของมันในสังคมสมัยใหม่นั้นกว้างมาก โรคจำนวนมาก รวมทั้งโรคที่แสดงออกในระดับพันธุกรรม กำหนดให้นักวิทยาศาสตร์ต้องหาทางแก้ไขปัญหานี้
คำอธิบายของกิจกรรม
ไวรัสและแบคทีเรียมีการกลายพันธุ์อย่างต่อเนื่อง ซึ่งหมายความว่ายาไม่ได้ช่วยให้คนและโรคต่างๆ รักษาไม่หายอีกต่อไป งานของอณูชีววิทยาคือการก้าวไปข้างหน้าของกระบวนการนี้และพัฒนาวิธีการรักษาโรคแบบใหม่ นักวิทยาศาสตร์ทำงานตามแผนที่กำหนดไว้อย่างดี: ปิดกั้นสาเหตุของโรค ขจัดกลไกการถ่ายทอดทางพันธุกรรม และด้วยเหตุนี้จึงบรรเทาสภาพของผู้ป่วย มีศูนย์ คลินิก และโรงพยาบาลหลายแห่งทั่วโลกที่นักชีววิทยาระดับโมเลกุลกำลังพัฒนาวิธีการรักษาแบบใหม่เพื่อช่วยเหลือผู้ป่วย
ความรับผิดชอบต่อหน้าที่
ความรับผิดชอบของนักชีววิทยาระดับโมเลกุลรวมถึงการศึกษากระบวนการภายในเซลล์ (เช่น การเปลี่ยนแปลงของ DNA ระหว่างการพัฒนาของเนื้องอก) นอกจากนี้ ผู้เชี่ยวชาญยังได้ศึกษาคุณสมบัติของดีเอ็นเอ ผลกระทบต่อสิ่งมีชีวิตทั้งหมด และเซลล์เดียว การศึกษาดังกล่าวดำเนินการบนพื้นฐานของ PCR (ปฏิกิริยาลูกโซ่โพลีเมอเรส) ซึ่งช่วยให้คุณวิเคราะห์ร่างกายสำหรับการติดเชื้อ โรคทางพันธุกรรม และกำหนดความสัมพันธ์ทางชีวภาพ
คุณสมบัติของการเติบโตของอาชีพ
อาชีพนักชีววิทยาระดับโมเลกุลมีแนวโน้มที่ดีในสาขานี้ และวันนี้ก็อ้างว่าเป็นอาชีพแรกในการจัดอันดับวิชาชีพแพทย์ในอนาคต อย่างไรก็ตาม นักชีววิทยาระดับโมเลกุลไม่จำเป็นต้องอยู่ในสาขานี้ตลอดเวลา หากมีความประสงค์จะเปลี่ยนอาชีพ ก็สามารถอบรมขึ้นใหม่ได้ในตำแหน่งผู้จัดการฝ่ายขายอุปกรณ์ห้องปฏิบัติการ เริ่มพัฒนาเครื่องมือสำหรับการศึกษาต่างๆ หรือเปิดธุรกิจของตนเอง