Molekylærbiolog. Anvendt molekylærbiologi
Molekylærbiologi har opplevd en periode med rask utvikling av egne forskningsmetoder, som nå skiller seg fra biokjemi. Disse inkluderer spesielt metoder for genteknologi, kloning, kunstig uttrykk og genknockout. Siden DNA er den materielle bæreren av genetisk informasjon, har molekylærbiologi blitt mye nærmere genetikk, og molekylær genetikk ble dannet i krysset, som både er en del av genetikk og molekylærbiologi. Akkurat som molekylærbiologi gjør omfattende bruk av virus som forskningsverktøy, bruker virologi metodene for molekylærbiologi for å løse sine problemer. Datateknologi er involvert i analyse av genetisk informasjon, i forbindelse med hvilke nye områder av molekylær genetikk har dukket opp, som noen ganger betraktes som spesielle disipliner: bioinformatikk, genomikk og proteomikk.
Utviklingshistorie
Denne banebrytende oppdagelsen ble utarbeidet av en lang fase med forskning på genetikk og biokjemi av virus og bakterier.
I 1928 viste Frederick Griffith først at et ekstrakt av varmedrepte patogene bakterier kunne overføre egenskapen patogenitet til godartede bakterier. Studiet av bakteriell transformasjon førte videre til rensing av sykdomsmidlet, som mot forventning viste seg å ikke være et protein, men en nukleinsyre. Selve nukleinsyren er ikke farlig, den bærer bare genene som bestemmer patogenisiteten og andre egenskaper til mikroorganismen.
På 50-tallet av XX-tallet ble det vist at bakterier har en primitiv seksuell prosess, de er i stand til å utveksle ekstrakromosomalt DNA, plasmider. Oppdagelsen av plasmider, så vel som transformasjoner, dannet grunnlaget for plasmidteknologien som er vanlig innen molekylærbiologi. En annen viktig oppdagelse for metodikken var oppdagelsen på begynnelsen av 1900-tallet av bakterielle virus, bakteriofager. Fager kan også overføre genetisk materiale fra en bakteriecelle til en annen. Infeksjon av bakterier av fager fører til en endring i sammensetningen av bakteriell RNA. Hvis sammensetningen av RNA uten fag ligner sammensetningen av bakteriell DNA, blir RNA etter infeksjon mer lik bakteriofag-DNA. Dermed ble det funnet at strukturen til RNA bestemmes av strukturen til DNA. I sin tur avhenger hastigheten av proteinsyntese i cellene av mengden av RNA-proteinkomplekser. Slik ble det formulert sentrale dogme innen molekylærbiologi: DNA ↔ RNA → protein.
Den videre utviklingen av molekylærbiologi ble ledsaget av både utviklingen av dens metodikk, spesielt oppfinnelsen av en metode for å bestemme nukleotidsekvensen til DNA (W. Gilbert og F. Sanger, Nobelprisen i kjemi i 1980), og nye funn innen forskning på geners struktur og funksjon (se. Genetikkens historie). Ved begynnelsen av det 21. århundre ble det innhentet data om den primære strukturen til alt menneskelig DNA og en rekke andre organismer, de viktigste for medisin, landbruk og vitenskapelig forskning, noe som førte til fremveksten av flere nye områder innen biologi: genomikk , bioinformatikk, etc.
se også
- Molekylærbiologi (tidsskrift)
- Transkriptomikk
- Molekylær paleontologi
- EMBO - European Organization for Molecular Biology
Litteratur
- Sanger M., Berg P. Gener og genomer. - Moskva, 1998.
- Stent G., Kalindar R. Molekylær genetikk. - Moskva, 1981.
- Sambrook J., Fritsch E.F., Maniatis T. Molekylær kloning. - 1989.
- Patrushev L.I. Uttrykk av gener. - M.: Nauka, 2000. - 000 s., ill. ISBN 5-02-001890-2
Lenker
Wikimedia Foundation. 2010 .
- Ardatovsky-distriktet i Nizhny Novgorod-regionen
- Arzamas-distriktet i Nizhny Novgorod-regionen
Se hva "Molecular Biology" er i andre ordbøker:
MOLEKYLBIOLOGI- studerer det grunnleggende. egenskaper og manifestasjoner av liv på molekylært nivå. De viktigste retningene i M. b. er studier av den strukturelle og funksjonelle organiseringen av det genetiske apparatet til celler og mekanismen for implementering av arvelig informasjon ... ... Biologisk leksikon ordbok
MOLEKYLBIOLOGI- utforsker livets grunnleggende egenskaper og manifestasjoner på molekylært nivå. Finner ut hvordan og i hvilken grad vekst og utvikling av organismer, lagring og overføring av arvelig informasjon, konvertering av energi i levende celler og andre fenomener skyldes ... Stor encyklopedisk ordbok
MOLEKYLBIOLOGI Moderne leksikon
MOLEKYLBIOLOGI- MOLEKYLÆR BIOLOGI, den biologiske studien av strukturen og funksjonen til MOLEKYLENE som utgjør levende organismer. De viktigste studieområdene inkluderer de fysiske og kjemiske egenskapene til proteiner og NUKLEINSYRER som DNA. se også… … Vitenskapelig og teknisk encyklopedisk ordbok
molekylbiologi- en del av biol., som utforsker de grunnleggende egenskapene og manifestasjonene av liv på molekylært nivå. Finner ut hvordan og i hvilken grad vekst og utvikling av organismer, lagring og overføring av arvelig informasjon, konvertering av energi i levende celler og ... ... Ordbok for mikrobiologi
molekylbiologi- — Emner innen bioteknologi EN molekylærbiologi … Teknisk oversetterhåndbok
Molekylbiologi- MOLEKYLÆR BIOLOGI, utforsker livets grunnleggende egenskaper og manifestasjoner på molekylært nivå. Finner ut hvordan og i hvilken grad vekst og utvikling av organismer, lagring og overføring av arvelig informasjon, konvertering av energi i levende celler og ... ... Illustrert encyklopedisk ordbok
Molekylbiologi- en vitenskap som setter som sin oppgave kunnskapen om livsfenomens natur ved å studere biologiske objekter og systemer på et nivå som nærmer seg det molekylære nivået, og i noen tilfeller når denne grensen. Sluttmålet med dette er…… Stor sovjetisk leksikon
MOLEKYLBIOLOGI- studerer livsfenomener på nivå med makromolekyler (kap. arr. proteiner og nukleinsyrer) i cellefrie strukturer (ribosomer, etc.), i virus, og også i celler. M.s formål. etablere rollen og mekanismen for funksjon av disse makromolekylene basert på ... ... Kjemisk leksikon
molekylbiologi- utforsker livets grunnleggende egenskaper og manifestasjoner på molekylært nivå. Finner ut hvordan og i hvilken grad vekst og utvikling av organismer, lagring og overføring av arvelig informasjon, konvertering av energi i levende celler og andre fenomener ... ... encyklopedisk ordbok
Bøker
- Cellens molekylærbiologi. Problembok, J. Wilson, T. Hunt. Boken til amerikanske forfattere er et vedlegg til 2. utgave av læreboken `Molecular Biology of the Cell` av B. Alberts, D. Bray, J. Lewis m.fl. Inneholder spørsmål og oppgaver, hvis formål er å utdype . ..
Molekylbiologi, en vitenskap som setter som sin oppgave kunnskapen om livsfenomens natur ved å studere biologiske objekter og systemer på et nivå som nærmer seg det molekylære nivået, og i noen tilfeller når denne grensen. Det endelige målet i dette tilfellet er å finne ut hvordan og i hvilken grad livets karakteristiske manifestasjoner, som arv, reproduksjon av ens egen art, proteinbiosyntese, eksitabilitet, vekst og utvikling, lagring og overføring av informasjon, energitransformasjoner, mobilitet , etc. , skyldes strukturen, egenskapene og interaksjonen til molekyler av biologisk viktige stoffer, først og fremst de to hovedklassene av høymolekylære biopolymerer - proteiner og nukleinsyrer. Et særtrekk ved M. b. - studiet av livets fenomener på livløse gjenstander eller de som er preget av livets mest primitive manifestasjoner. Dette er biologiske formasjoner fra cellenivå og nedenfor: subcellulære organeller, som isolerte cellekjerner, mitokondrier, ribosomer, kromosomer, cellemembraner; videre - systemer som står på grensen mellom levende og livløs natur - virus, inkludert bakteriofager, og slutter med molekylene til de viktigste komponentene i levende materie - nukleinsyrer og proteiner.
Grunnlaget som M. utviklet seg på ble lagt av slike vitenskaper som genetikk, biokjemi, fysiologi av elementære prosesser, etc. I henhold til opprinnelsen til utviklingen, M. b. er uløselig knyttet til molekylær genetikk, som fortsatt er en viktig del av
Et særtrekk ved M. b. er dens tredimensjonalitet. Essensen av M. b. er sett av M. Perutz i å tolke biologiske funksjoner i form av molekylær struktur. M. b. har som mål å få svar på spørsmålet "hvordan", å vite essensen av rollen og deltakelsen til hele strukturen til molekylet, og på spørsmålene "hvorfor" og "hva for", å finne ut på den ene siden forholdet mellom egenskapene til molekylet (igjen, primært proteiner og nukleinsyrer) og funksjonene det utfører, og på den annen side rollen til slike individuelle funksjoner i det totale komplekset av manifestasjoner av vital aktivitet.
De viktigste prestasjonene innen molekylærbiologi. Her er en langt fra fullstendig liste over disse prestasjonene: avsløring av strukturen og mekanismen for den biologiske funksjonen til DNA, alle typer RNA og ribosomer, avsløring av den genetiske koden; oppdagelse av revers transkripsjon, dvs. DNA-syntese på en RNA-mal; studie av mekanismene for funksjon av respiratoriske pigmenter; oppdagelse av den tredimensjonale strukturen og dens funksjonelle rolle i virkningen av enzymer, prinsippet om matrisesyntese og mekanismene for proteinbiosyntese; avsløring av strukturen til virus og mekanismene for deres replikasjon, den primære og, delvis, den romlige strukturen til antistoffer; isolering av individuelle gener, kjemisk og deretter biologisk (enzymatisk) gensyntese, inkludert menneskelig, utenfor cellen (in vitro); overføring av gener fra en organisme til en annen, inkludert til menneskelige celler; den raskt fremadskridende dechiffreringen av den kjemiske strukturen til et økende antall individuelle proteiner, hovedsakelig enzymer, så vel som nukleinsyrer; oppdagelse av fenomenene "selvmontering" av noen biologiske objekter med stadig økende kompleksitet, med utgangspunkt i nukleinsyremolekyler og videre til flerkomponentenzymer, virus, ribosomer, etc.; belysning av allosteriske og andre grunnleggende prinsipper for regulering av biologiske funksjoner og prosesser.
Problemer med molekylærbiologi. Sammen med de angitte viktige oppgaver ville M.. (kunnskap om lovene om "anerkjennelse", selvmontering og integrasjon) en faktisk retning for vitenskapelig søken etter nær fremtid er utviklingen av metoder som tillater dechiffrering av strukturen, og deretter den tredimensjonale, romlige organiseringen av høymolekylære nukleinsyrer. Alle de viktigste metodene, hvis bruk sikret fremveksten og suksessen til M. b., ble foreslått og utviklet av fysikere (ultrasentrifugering, røntgendiffraksjonsanalyse, elektronmikroskopi, kjernemagnetisk resonans, etc.). Nesten alle nye fysiske eksperimentelle tilnærminger (for eksempel bruk av datamaskiner, synkrotron eller bremsstrahlung, stråling, laserteknologi og andre) åpner for nye muligheter for en dyptgående studie av problemene med meteorologisk analyse. Blant de viktigste oppgavene av praktisk art, svaret som forventes fra M. b., er i første omgang problemet med det molekylære grunnlaget for ondartet vekst, deretter - måter å forebygge og kanskje overvinne arvelige sykdommer - " molekylære sykdommer". Av stor betydning vil være belysningen av det molekylære grunnlaget for biologisk katalyse, dvs. virkningen av enzymer. Blant de viktigste moderne retningene til M. b. bør inkludere ønsket om å dechiffrere de molekylære virkningsmekanismene til hormoner, giftige og medisinske stoffer, samt å finne ut detaljene i den molekylære strukturen og funksjonen til slike cellulære strukturer som biologiske membraner involvert i reguleringen av penetrasjonsprosessene og transport av stoffer. Fjernere mål M. b. - kunnskap om arten av nerveprosesser, hukommelsesmekanismer, etc. En av de viktige nye delene av M. b. - såkalte. genteknologi, som tar sikte på målrettet å betjene det genetiske apparatet (genomet) til levende organismer, starter med mikrober og lavere (encellet) og slutter med mennesker (i sistnevnte tilfelle, primært med det formål å radikal behandling av arvelige sykdommer og korrigering av genetiske defekter).
De viktigste retningene til MB:
- Molekylær genetikk - studiet av den strukturelle og funksjonelle organiseringen av det genetiske apparatet til cellen og mekanismen for implementering av arvelig informasjon
– Molekylær virologi – studiet av de molekylære mekanismene for interaksjonen mellom virus og celler
– Molekylær immunologi – studiet av mønstre av immunreaksjoner i kroppen
- Molekylær biologi for utvikling - studiet av utseendet til cellemangfold i løpet av individuell utvikling av organismer og spesialisering av celler
Hovedformål med forskning: Virus (inkludert bakteriofager), Celler og subcellulære strukturer, Makromolekyler, Flercellede organismer.
Utviklingen av biokjemi, biofysikk, genetikk, cytokjemi, mange deler av mikrobiologi og virologi rundt begynnelsen av 40-tallet av XX-tallet. tett ført til studiet av livsfenomener på molekylært nivå. Suksessene oppnådd av disse vitenskapene, samtidig og fra forskjellige sider, førte til erkjennelsen av det faktum at det er på molekylært nivå at kroppens hovedkontrollsystemer fungerer, og at den videre fremgangen til disse vitenskapene vil avhenge av avsløringen av de biologiske funksjonene til molekylene som utgjør organismenes kropper, deres deltakelse i syntese og desintegrasjon, gjensidige transformasjoner og reproduksjon av forbindelser i cellen, samt utveksling av energi og informasjon som skjer i dette tilfellet. Dermed oppsto en helt ny gren - molekylærbiologien, i skjæringspunktet mellom disse biologiske disiplinene med kjemi og fysikk.
I motsetning til biokjemi, er oppmerksomheten til moderne molekylærbiologi hovedsakelig fokusert på studiet av strukturen og funksjonen til de viktigste klassene av biopolymerer - proteiner og nukleinsyrer, hvorav den første bestemmer selve muligheten for metabolske reaksjoner, og den andre - den biosyntese av spesifikke proteiner. Det er derfor klart at det er umulig å gjøre et klart skille mellom molekylærbiologi og biokjemi, de tilsvarende grenene av genetikk, mikrobiologi og virologi.
Fremveksten av molekylærbiologi var nært knyttet til utviklingen av nye forskningsmetoder, som allerede er diskutert i de aktuelle kapitlene. Sammen med utviklingen av elektronmikroskopi og andre metoder for mikroskopisk teknikk, spilte metodene for fraksjonering av cellulære elementer utviklet på 1950-tallet en viktig rolle. De var basert på forbedrede metoder for differensiell sentrifugering (A. Claude, 1954). På dette tidspunktet var det allerede ganske pålitelige metoder for isolering og fraksjonering av biopolymerer. Dette inkluderer spesielt metoden for proteinfraksjonering ved elektroforese foreslått av A. Tiselius (1937; Nobelprisen, 1948), metoder for isolering og rensing av nukleinsyrer (E. Kay, A. Downs, M. Sevag, A. Mirsky , og andre. ). Samtidig ble forskjellige metoder for kromatografisk analyse utviklet i mange laboratorier i verden (A. Martin og R. Sing, 1941; Nobelprisen, 1952), og ble deretter betydelig forbedret.
Røntgendiffraksjonsanalyse spilte en uvurderlig tjeneste ved å dechiffrere strukturen til biopolymerer. De grunnleggende prinsippene for røntgendiffraksjonsanalyse ble utviklet ved King's College London University under ledelse av W. Bragg av en gruppe forskere, som inkluderte J. Bernal, A. Londsdale, W. Astbury, J. Robertson og andre.
Spesielt bør nevnes studiene av Protoplasm Biochemistry (1925 - 1929), professor ved Moscow State University A.R. Kizel, som var av stor betydning for den etterfølgende utviklingen av molekylærbiologi. Kizel ga et slag mot den fast forankrede forestillingen om at enhver protoplasma er basert på en spesiell proteinkropp - plater, som angivelig bestemmer alle dens viktigste strukturelle og funksjonelle egenskaper. Han viste at plater er et protein som bare finnes i myxomyceter, og da på et visst utviklingsstadium, og at ingen permanent komponent - et enkelt skjelettprotein - finnes i protoplasma. Dermed tok studiet av problemet med strukturen til protoplasma og den funksjonelle rollen til proteiner den rette veien og fikk rom for utviklingen. Kisels forskning har vunnet verdensomspennende anerkjennelse, og stimulerer studiet av kjemien til cellens bestanddeler.
Begrepet «molekylær biologi», først brukt av den engelske krystallografen W. Astbury, professor ved University of Leeds, dukket sannsynligvis opp tidlig på 1940-tallet (før 1945). De grunnleggende røntgendiffraksjonsstudiene av proteiner og DNA, utført av Astbury på 1930-tallet, tjente som grunnlag for den påfølgende vellykkede dechiffreringen av den sekundære strukturen til disse biopolymerene. I 1963 skrev J. Bernal: "Et monument over ham vil bli reist av hele molekylærbiologien - vitenskapen som han ga navn og virkelig grunnla" * , I litteraturen dukket dette begrepet opp for første gang, kanskje i 1946 i artikkelen av W. Astbury "Progress in X-ray diffraction analysis of organic and fibrillar compounds", publisert i det engelske tidsskriftet "Nature" ** . I sin Harvey-forelesning bemerket Astbury (1950): "Jeg er glad for at begrepet molekylærbiologi nå er ganske mye brukt, selv om det er usannsynlig at jeg var den første som foreslo det. Jeg likte det og jeg har lenge prøvd å spre det. "***. Allerede i 1950 var Astbury tydelig på at molekylærbiologi først og fremst omhandler strukturen og konformasjonen til makromolekyler, hvor studiet er av avgjørende betydning for å forstå hvordan levende organismer fungerer.
* (biogr. Mem. Felles Roy. Soc, 1963, v. 9, 29.)
** (W.T. Astbury. Fremdrift av røntgenanalyse av organiske og fiberstrukturer.- Nature,. 1946, v. 157, 121.)
*** (W.T. Astbury. Eventyr i molekylærbiologi. Thomas Springfield, 1952, s. 3.)
Molekylærbiologi har møtt og står faktisk overfor de samme oppgavene som biologien som helhet - kunnskapen om livets essens og dets hovedfenomener, spesielt, som arv og variabilitet. Moderne molekylærbiologi er først og fremst ment å dechiffrere strukturen og funksjonen til gener, måtene og mekanismene for realisering av den genetiske informasjonen til organismer på forskjellige stadier av ontogenese og på forskjellige stadier av lesingen. Den er designet for å avsløre de subtile mekanismene for regulering av genaktivitet og celledifferensiering, for å belyse arten av mutagenese og det molekylære grunnlaget for den evolusjonære prosessen.
Etablering av den genetiske rollen til nukleinsyrer
For utviklingen av molekylærbiologi var følgende funn av største betydning. I 1944 viste amerikanske forskere O. Avery, K. McLeod (Nobelprisen, 1923) og M. McCarthy at DNA-molekyler isolert fra pneumokokker har transformerende aktivitet. Etter hydrolyse av disse DNA-ene med deoksyribonuklease, forsvant deres transformerende aktivitet fullstendig. Dermed ble det for første gang overbevisende bevist at det er DNA, og ikke protein, som er utstyrt med genetiske funksjoner i en celle.
I rettferdighet bør det bemerkes at fenomenet bakteriell transformasjon ble oppdaget mye tidligere enn oppdagelsen av Avery, McLeod og McCarthy. I 1928 publiserte F. Griffith en artikkel der han rapporterte at etter tilsetning av drepte celler av en innkapslet virulent stamme til ikke-virulente (ikke-innkapslede) pneumokokker, blir den resulterende blandingen av celler dødelig for mus. Videre var levende pneumokokkceller isolert fra dyr infisert med denne blandingen allerede virulente og hadde en polysakkaridkapsel. I dette eksperimentet ble det således vist at under påvirkning av enkelte komponenter i de drepte pneumokokkcellene, blir den ikke-innkapslede bakterieformen til en kapseldannende virulent form. Seksten år senere erstattet Avery, McLeod og McCarthy drepte hele pneumokokkceller med deres deoksyribonukleinsyre i dette eksperimentet og viste at det var DNA som hadde transformerende aktivitet (se også kapittel 7 og 25). Betydningen av denne oppdagelsen er vanskelig å overvurdere. Det stimulerte studiet av nukleinsyrer i mange laboratorier rundt om i verden og tvang forskere til å fokusere på DNA.
Sammen med oppdagelsen av Avery, McLeod og McCarthy, på begynnelsen av 1950-tallet, hadde en ganske stor mengde direkte og indirekte bevis allerede samlet seg på at nukleinsyrer spiller en eksepsjonell rolle i livet og har en genetisk funksjon. Dette ble spesielt indikert av naturen til DNA-lokalisering i cellen og dataene til R. Vendrelli (1948) at DNA-innholdet per celle er strengt konstant og korrelerer med graden av ploiditet: i haploide kjønnsceller er DNA halvparten av diploide somatiske celler. Den uttalte metabolske stabiliteten til DNA vitnet også til fordel for den genetiske rollen til DNA. På begynnelsen av 50-tallet hadde mange forskjellige fakta samlet seg, noe som tyder på at de fleste kjente mutagene faktorene virker hovedsakelig på nukleinsyrer og spesielt på DNA (R. Hotchkiss, 1949; G. Ephrussi-Taylor, 1951; E. Freese, 1957 og andre).
Av spesiell betydning for å etablere den genetiske rollen til nukleinsyrer var studiet av ulike fager og virus. I 1933 fant D. Schlesinger DNA i bakteriofagen til Escherichia coli. Siden isoleringen av tobakksmosaikkvirus (TMV) i krystallinsk tilstand av W. Stanley (1935, Nobelprisen, 1946), har et nytt stadium i studiet av plantevirus begynt. I 1937 - 1938. ansatte ved Rothamsted Agricultural Station (England) F. Bowden og N. Pirie viste at mange plantevirus isolert av dem ikke er globuliner, men er ribonukleoproteiner og inneholder nukleinsyre som en obligatorisk komponent. Helt på begynnelsen av 40-tallet ble verkene til G. Schramm (1940), PA Agatov (1941), G. Miller og W. Stanley (1941) publisert, noe som indikerer at en merkbar kjemisk modifikasjon av proteinkomponenten ikke fører til til tap av TMV-infeksjon. Dette indikerte at proteinkomponenten ikke kunne være bærer av virusets arvelige egenskaper, slik mange mikrobiologer fortsatte å tro. Overbevisende bevis til fordel for den genetiske rollen til nukleinsyre (RNA) i plantevirus ble oppnådd i 1956 av G. Schramm i Tübingen (FRG) og H. Frenkel-Konrath i California (USA). Disse forskerne nesten samtidig og uavhengig av hverandre isolerte RNA fra TMV og viste at det, og ikke proteinet, har smitteevne: som et resultat av infeksjon av tobakksplanter med dette RNA, ble normale virale partikler dannet og multiplisert i dem. Dette betydde at RNA inneholdt informasjon for syntese og sammenstilling av alle virale komponenter, inkludert virusproteinet. I 1968 slo I. G. Atabekov fast at protein spiller en betydelig rolle i selve infeksjonen av planter - proteinets natur bestemmer spekteret til vertsplanter.
I 1957 utførte Frenkel-Konrat for første gang rekonstruksjonen av TMV fra dens bestanddeler - RNA og protein. Sammen med normale partikler fikk han blandede "hybrider" der RNA var fra en stamme og proteinet fra en annen. Arveligheten til slike hybrider ble fullstendig bestemt av RNA, og avkom av virusene tilhørte stammen hvis RNA ble brukt for å oppnå de første blandede partiklene. Senere viste eksperimentene til A. Gierer, G. Schuster og G. Schramm (1958) og G. Witman (1960 - 1966) at den kjemiske modifikasjonen av TMV-nukleinkomponenten fører til oppkomsten av forskjellige mutanter av dette viruset.
I 1970 fant D. Baltimore og G. Temin at overføring av genetisk informasjon ikke bare kan skje fra DNA til RNA, men omvendt. De fant i noen onkogene RNA-holdige virus (oncornavirus) et spesielt enzym, den såkalte revers transkriptase, som er i stand til å syntetisere komplementært DNA på RNA-kjeder. Denne store oppdagelsen gjorde det mulig å forstå mekanismen for innsetting av den genetiske informasjonen til RNA-holdige virus i vertsgenomet og å ta en ny titt på arten av deres onkogene virkning.
Oppdagelse av nukleinsyrer og studie av deres egenskaper
Begrepet nukleinsyrer ble introdusert av den tyske biokjemikeren R. Altman i 1889, etter at disse forbindelsene ble oppdaget i 1869 av den sveitsiske legen F. Miescher. Misher ekstraherte puscellene med fortynnet saltsyre i flere uker og oppnådde nesten rent kjernefysisk materiale i resten. Han anså dette materialet for å være et karakteristisk "stoff av cellekjerner og kalte det nuklein. Når det gjelder egenskaper, skilte nuklein seg kraftig fra proteiner: det var surere, inneholdt ikke svovel, men det inneholdt mye fosfor, det var lett løselig i alkalier, men løste seg ikke opp i fortynnede syrer.
Misher sendte resultatene av sine observasjoner på nuklein til F. Goppe-Seyler for publisering i et tidsskrift. Stoffet han beskrev var så uvanlig (på det tidspunktet var det kun lecitin som var kjent av alle biologiske fosforholdige forbindelser) at Goppe-Seyler ikke trodde på Mishers eksperimenter, returnerte manuskriptet til ham og instruerte sine ansatte N. Plosh og N. Lyubavin til å sjekk konklusjonene hans på annet materiale. Mieschers verk "Om den kjemiske sammensetningen av pusceller" ble utgitt to år senere (1871). Samtidig ble verkene til Goppe-Seyler og hans samarbeidspartnere publisert om sammensetningen av pusceller, erytrocytter av fugler, slanger og andre celler. I løpet av de neste tre årene ble nuklein isolert fra dyreceller og gjær.
I sitt arbeid bemerket Misher at en detaljert studie av forskjellige nukleiner kan føre til etablering av forskjeller mellom dem, og dermed forutse ideen om spesifisitet til nukleinsyrer. Mens han studerte laksemelk, fant Misher at nukleinet i dem er i form av salt og er assosiert med hovedproteinet, som han kalte protamin.
I 1879 begynte A. Kossel å studere nukleiner i laboratoriet til Goppe-Seyler. I 1881 isolerte han hypoxanthin fra nuklein, men på den tiden tvilte han fortsatt på opprinnelsen til denne basen og trodde at hypoxanthin kunne være et produkt av proteinnedbrytning. I 1891, blant produktene fra nukleinhydrolyse, oppdaget Kossel adenin, guanin, fosforsyre og et annet stoff med egenskapene til sukker. For forskning på kjemien til nukleinsyrer ble Kossel tildelt Nobelprisen i 1910.
Ytterligere fremgang i å dechiffrere strukturen til nukleinsyrer er assosiert med forskningen til P. Levin og kolleger (1911 - 1934). I 1911 identifiserte P. Levin og V. Jacobs karbohydratkomponenten i adenosin og guanosin; de fant ut at disse nukleosidene inneholder D-ribose. I 1930 viste Lewin at karbohydratkomponenten i deoksyribonukleosider er 2-deoksy-D-ribose. Fra hans arbeid ble det kjent at nukleinsyrer bygges fra nukleotider, dvs. fosforylerte nukleosider. Levin mente at hovedtypen binding i nukleinsyrer (RNA) er 2", 5" fosfodiesterbindingen. Denne oppfatningen viste seg å være feil. Takket være arbeidet til den engelske kjemikeren A. Todd (Nobelprisen, 1957) og hans samarbeidspartnere, samt de engelske biokjemikerne R. Markham og J. Smith, ble det tidlig på 50-tallet kjent at hovedtypen binding i RNA er 3", 5" - fosfodiesterbinding.
Lewin viste at forskjellige nukleinsyrer kan variere i karbohydratkomponentens natur: noen av dem inneholder sukkeret deoksyribose, mens andre inneholder ribose. I tillegg skilte disse to typene nukleinsyrer seg i naturen til en av basene: pentose-type nukleinsyrer inneholdt uracil, og deoksypentose-type nukleinsyrer inneholdt tymin. Deoksypentosenukleinsyre (i moderne terminologi, deoksyribonukleinsyre - DNA) ble vanligvis lett isolert i store mengder fra thymus (søt kjertel) hos kalver. Derfor ble det kalt tymonukleinsyre. Kilden til pentose-type nukleinsyre (RNA) var hovedsakelig gjær og hvetekim. Denne typen ble ofte referert til som gjærnukleinsyre.
På begynnelsen av 1930-tallet var forestillingen om at planteceller var preget av en gjær-type nukleinsyre ganske fast forankret, mens tymonukleinsyre kun var karakteristisk for kjernene til dyreceller. De to typene nukleinsyrer, RNA og DNA, ble da kalt henholdsvis plante- og dyrenukleinsyrer. Imidlertid, som de tidlige studiene av A. N. Belozersky viste, er en slik deling av nukleinsyrer uberettiget. I 1934 oppdaget Belozersky først tymonukleinsyre i planteceller: fra erteplanter isolerte og identifiserte han tymin-pyrimidinbasen, som er karakteristisk for DNA. Så oppdaget han tymin i andre planter (soyafrø, bønner). I 1936 isolerte A. N. Belozersky og I. I. Dubrovskaya DNA forberedende fra hestekastanjefrøplanter. I tillegg viste en serie studier utført i England på 1940-tallet av D. Davidson og medarbeidere på en overbevisende måte at plantenukleinsyre (RNA) finnes i mange dyreceller.
Den utbredte bruken av den cytokjemiske reaksjonen for DNA utviklet av R. Felgen og G. Rosenbeck (1924) og reaksjonen til J. Brachet (1944) for RNA gjorde det mulig å raskt og entydig løse spørsmålet om den foretrukne lokaliseringen av disse nukleinene syrer i cellen. Det viste seg at DNA er konsentrert i kjernen, mens RNA overveiende er konsentrert i cytoplasma. Senere ble det funnet at RNA finnes både i cytoplasma og i kjernen, og i tillegg ble cytoplasmatisk DNA identifisert.
Når det gjelder spørsmålet om den primære strukturen til nukleinsyrer, på midten av 1940-tallet, var P. Levins idé solid etablert i vitenskapen, ifølge hvilken alle nukleinsyrer er bygget etter samme type og består av det samme såkalte tetranukleotidet. blokker. Hver av disse blokkene, ifølge Lewin, inneholder fire forskjellige nukleotider. Tetranukleotidteorien om strukturen til nukleinsyrer fratok i stor grad disse biopolymerene for spesifisitet. Derfor er det ikke overraskende at på den tiden var alle spesifikasjonene til levende ting bare assosiert med proteiner, hvis art av monomerer er mye mer mangfoldig (20 aminosyrer).
Det første gapet i teorien om tetranukleotidstrukturen til nukleinsyrer ble laget av analytiske data fra den engelske kjemikeren J. Gouland (1945 - 1947). Ved å bestemme sammensetningen av nukleinsyrer ved basenitrogenet, oppnådde han ikke et ekvimolart forhold av baser, slik det burde vært ifølge Lewins teori. Til slutt kollapset tetranukleotidteorien om strukturen til nukleinsyrer som et resultat av forskningen til E. Chargaff og hans samarbeidspartnere (1949 - 1951). Chargaff brukte papirkromatografi for å separere basene som ble frigjort fra DNA som et resultat av syrehydrolyse. Hver av disse basene ble nøyaktig bestemt spektrofotometrisk. Chargaff la merke til betydelige avvik fra det ekvimolare forholdet mellom baser i DNA av forskjellig opprinnelse og uttalte for første gang definitivt at DNA har en uttalt artsspesifisitet. Dette avsluttet hegemoniet til konseptet proteinspesifisitet i den levende cellen. Ved å analysere DNA av forskjellig opprinnelse, oppdaget og formulerte Chargaff unike mønstre av DNA-sammensetning, som kom inn i vitenskapen under navnet Chargaffs regler. I henhold til disse reglene, i alt DNA, uavhengig av opprinnelse, er mengden adenin lik mengden tymin (A = T), mengden guanin er lik mengden cytosin (G = C), mengden av puriner er lik mengden pyrimidiner (G + A = C + T), mengden baser med 6-aminogrupper er lik antall baser med 6-ketogrupper (A + C = G + T). Samtidig, til tross for slike strenge kvantitative samsvar, er DNA fra forskjellige arter forskjellig i verdien av A+T:G+C-forholdet. I noen DNA er det mengden guanin og cytosin som råder over mengden adenin og tymin (Chargaff kalte disse DNA GC-type DNA); andre DNA-er inneholdt mer adenin og tymin enn guanin og cytosin (disse DNA-ene ble kalt AT-type DNA). Dataene innhentet av Chargaff om sammensetningen av DNA spilte en eksepsjonell rolle i molekylærbiologi. Det var de som dannet grunnlaget for oppdagelsen av strukturen til DNA, laget i 1953 av J. Watson og F. Crick.
Tilbake i 1938 viste W. Astbury og F. Bell, ved hjelp av røntgendiffraksjonsanalyse, at baseplanene i DNA skulle være vinkelrett på molekylets langakse og så å si ligne en stabel med plater som ligger en over. den andre. Med forbedring av teknikken for røntgendiffraksjonsanalyse, innen 1952 - 1953. akkumulert informasjon som gjorde det mulig å bedømme lengden på individuelle bindinger og helningsvinklene. Dette gjorde det mulig å representere med størst sannsynlighet arten av orienteringen til ringene av pentoserester i sukkerfosfat-ryggraden til DNA-molekylet. I 1952 foreslo S. Farberg to spekulative modeller av DNA, som representerte et enkelttrådet molekyl foldet eller vridd på seg selv. En ikke mindre spekulativ modell av strukturen til DNA ble foreslått i 1953 av L. Pauling (Nobelprisvinner, 1954) og R. Corey. I denne modellen dannet tre vridd DNA-tråder en lang helix, hvis kjerne var representert av fosfatgrupper, og basene var plassert utenfor den. I 1953 oppnådde M. Wilkins og R. Franklin klarere røntgendiffraksjonsmønstre av DNA. Analysen deres viste fullstendig feil i modellene til Farberg, Pauling og Corey. Ved å bruke Chargaffs data, sammenligne ulike kombinasjoner av molekylære modeller av individuelle monomerer og røntgendiffraksjonsdata, kom J. Watson og F. Crick i 1953 til den konklusjon at DNA-molekylet må være en dobbelttrådet helix. Chargaffs regler begrenset sterkt antallet mulige ordnede kombinasjoner av baser i den foreslåtte DNA-modellen; de foreslo for Watson og Crick at det måtte være en spesifikk baseparing i DNA-molekylet - adenin med tymin, og guanin med cytosin. Med andre ord, adenin i en DNA-streng tilsvarer alltid strengt tatt tymin i den andre strengen, og guanin i en streng tilsvarer nødvendigvis cytosin i den andre. Dermed formulerte Watson og Crick for første gang prinsippet om den komplementære strukturen til DNA av eksepsjonell betydning, i henhold til hvilket en DNA-tråd komplementerer en annen, dvs. at basesekvensen til en streng unikt bestemmer sekvensen av baser i den andre (komplementært). ) Strand. Det ble åpenbart at allerede i selve strukturen til DNA ligger potensialet for dets eksakte reproduksjon. Denne modellen for DNA-struktur er for tiden generelt akseptert. Crick, Watson og Wilkins ble tildelt Nobelprisen i 1962 for å dechiffrere strukturen til DNA.
Det skal bemerkes at ideen om en mekanisme for nøyaktig reproduksjon av makromolekyler og overføring av arvelig informasjon oppsto i vårt land. I 1927 foreslo N. K. Koltsov at under cellereproduksjon skjer reproduksjonen av molekyler ved eksakt autokatalytisk reproduksjon av de eksisterende modermolekylene. Riktignok ga Koltsov på den tiden denne egenskapen ikke med DNA-molekyler, men med molekyler av proteinnatur, hvis funksjonelle betydning da var ukjent. Ikke desto mindre viste selve ideen om autokatalytisk reproduksjon av makromolekyler og mekanismen for overføring av arvelige egenskaper å være profetisk: det ble den ledende ideen om moderne molekylærbiologi.
Utført i laboratoriet til A. N. Belozersky av A. S. Spirin, G. N. Zaitseva, B. F. Vanyushin, S. O. Uryson, A. S. Antonov og andre forskjellige organismer bekreftet fullt ut mønstrene oppdaget av Chargaff, og full samsvar med den molekylære modellen for DNA-strukturen foreslått av Watson og Crick. Disse studiene har vist at DNA fra forskjellige bakterier, sopp, alger, actinomycetes, høyere planter, virvelløse dyr og virveldyr har en spesifikk sammensetning. Forskjeller i sammensetningen (innholdet av AT-basepar) er spesielt uttalt i mikroorganismer, og viser seg å være et viktig taksonomisk trekk. Hos høyere planter og dyr er artsvariasjoner i DNA-sammensetningen mye mindre uttalte. Men dette betyr ikke at deres DNA er mindre spesifikt. I tillegg til sammensetningen av baser, bestemmes spesifisiteten i stor grad av deres sekvens i DNA-kjeder.
Sammen med de vanlige basene ble det funnet ytterligere nitrogenholdige baser i DNA og RNA. G. White (1950) fant således 5-metylcytosin i DNA fra planter og dyr, og D. Dunn og J. Smith (1958) fant metylert adenin i noe DNA. I lang tid ble metylcytosin ansett som et kjennetegn på arvestoffet til høyere organismer. I 1968 fant A. N. Belozersky, B. F. Vanyushin og N. A. Kokurina ut at det også kan finnes i DNA til bakterier.
I 1964 oppdaget M. Gold og J. Hurwitz en ny klasse enzymer som utfører den naturlige modifikasjonen av DNA - dets metylering. Etter denne oppdagelsen ble det klart at mindre (inneholdt i små mengder) baser oppstår allerede på den ferdige DNA-polynukleotidkjeden som et resultat av spesifikk metylering av cytosin- og adeninrester i spesielle sekvenser. Spesielt, ifølge B. F. Vanyushin, Ya. I. Buryanov og A. N. Belozersky (1969), kan adeninmetylering i E. coli DNA forekomme i terminerende kodoner. I følge AN Belozersky og medarbeidere (1968 - 1970), samt M. Meselson (USA) og V. Arber (Sveits) (1965 - 1969), gir metylering unike individuelle trekk til DNA-molekyler og, i kombinasjon med virkningen av spesifikke nukleaser, er en del av en kompleks mekanisme som kontrollerer syntesen av DNA i cellen. Med andre ord, naturen til metylering av et bestemt DNA forhåndsbestemmer spørsmålet om det kan formere seg i en gitt celle.
Nesten samtidig startet isolasjonen og den intensive studien av DNA-metylaser og restriksjonsendonukleaser; i 1969 - 1975 nukleotidsekvensene gjenkjent i DNA av noen av disse enzymene er etablert (X. Boyer, X. Smith, S. Lynn, K. Murray). Når forskjellige DNA-er hydrolyseres av et restriksjonsenzym, blir ganske store fragmenter med identiske "klebrige" ender spaltet ut. Dette gjør det mulig ikke bare å analysere strukturen til gener, slik man gjør i små virus (D. Nathans, S. Adler, 1973 - 1975), men også å konstruere ulike genomer. Med oppdagelsen av disse spesifikke restriksjonsenzymene har genteknologi blitt en håndgripelig realitet. Innebygd i små plasmid-DNA-gener av forskjellig opprinnelse er allerede lett introdusert i forskjellige celler. Så en ny type biologisk aktive plasmider ble oppnådd som ga resistens mot visse antibiotika (S. Cohen, 1973), ribosomale gener fra en frosk og Drosophila ble introdusert i Escherichia coli-plasmider (J. Morrow, 1974; X. Boyer, D. Hogness, R. Davis, 1974 - 1975). Dermed er virkelige måter åpne for å skaffe fundamentalt nye organismer ved å introdusere og integrere ulike gener i deres genpool. Denne oppdagelsen kan rettes til fordel for hele menneskeheten.
I 1952 oppdaget G. White og S. Cohen at DNA-et til T-even fager inneholder en uvanlig base - 5-hydroksymetylcytosin. Senere, fra verkene til E. Volkin og R. Sinsheimer (1954) og Cohen (1956), ble det kjent at hydroksymetylcytosinrester kan bli fullstendig eller delvis glukosidisert, som et resultat av at fag-DNA-molekylet er beskyttet mot den hydrolytiske virkningen av nukleaser.
På begynnelsen av 1950-tallet, fra verkene til D. Dunn og J. Smith (England), S. Zamenhof (USA) og A. Wacker (Tyskland), ble det kjent at mange kunstige baseanaloger kan inkluderes i DNA, noen ganger erstatter de opptil 50 % tymin. Som regel fører disse substitusjonene til feil i DNA-replikasjon, transkripsjon og translasjon og til opptreden av mutanter. Således fant J. Marmur (1962) at DNA fra noen fager inneholder oksymetyluracil i stedet for tymin. I 1963 oppdaget I. Takahashi og J. Marmur at DNAet til en av fagene inneholder uracil i stedet for tymin. Dermed kollapset et annet prinsipp, ifølge hvilket nukleinsyrer tidligere ble separert. Siden tiden for P. Levins arbeid har det vært antatt at tymin er kjennetegnet for DNA, og uracil er kjennetegnet for RNA. Det ble klart at dette tegnet ikke alltid er pålitelig, og den grunnleggende forskjellen i den kjemiske naturen til de to typene nukleinsyrer, slik det ser ut i dag, er bare naturen til karbohydratkomponenten.
I studiet av fager har mange uvanlige trekk ved organiseringen av nukleinsyrer blitt avdekket. Siden 1953 har det vært antatt at alt DNA er dobbelttrådet lineære molekyler, mens RNA kun er enkelttrådet. Denne posisjonen ble betydelig rystet i 1961, da R. Sinsheimer oppdaget at DNAet til fagen φ X 174 er representert av et enkelttrådet sirkulært molekyl. Senere viste det seg imidlertid at i denne formen eksisterer dette DNA bare i en vegetativ fagpartikkel, og den replikative formen av DNAet til denne fagen er også dobbelttrådet. I tillegg viste det seg å være ganske uventet at RNA til enkelte virus kan være dobbelttrådet. Denne nye typen makromolekylær organisering av RNA ble oppdaget i 1962 av P. Gomatos, I. Tamm og andre forskere innen noen dyrevirus og i plantesårtumorvirus. Nylig fastslo V. I. Agol og A. A. Bogdanov (1970) at i tillegg til lineære RNA-molekyler, finnes det også lukkede eller sykliske molekyler. De oppdaget syklisk dobbelttrådet RNA, spesielt i encefalomyelokardittviruset. Takket være verkene til X. Deveaux, L. Tinoko, T. I. Tikhonenko, E. I. Budovsky og andre (1960 - 1974), ble hovedtrekkene i organiseringen (leggingen) av genetisk materiale i bakteriofager kjent.
På slutten av 1950-tallet fant den amerikanske forskeren P. Doty at oppvarming forårsaker DNA-denaturering, som er ledsaget av brudd av hydrogenbindinger mellom basepar og separasjon av komplementære kjeder. Denne prosessen har karakter av en "spiral-coil" faseovergang og ligner smelting av krystaller. Derfor kalte Doty prosessen med termisk denaturering av DNA-DNA-smelting. Med langsom avkjøling skjer renaturering av molekyler, dvs. gjenforening av komplementære halvdeler.
Prinsippet for renaturering i 1960 ble brukt av J. Marmur og K. Schildkraut for å bestemme graden av "hybridisering" av DNA fra forskjellige mikroorganismer. Deretter forbedret E. Bolton og B. McCarthy denne teknikken ved å foreslå metoden for de såkalte DNA-agar-søylene. Denne metoden viste seg å være uunnværlig for å studere graden av homologi til nukleotidsekvensen til forskjellige DNA og belyse det genetiske forholdet til forskjellige organismer. Denatureringen av DNA oppdaget av Doty i kombinasjon med kromatografi på metylert albumin beskrevet av J. Mandel og A. Hershey * (1960) og tetthetsgradientsentrifugering (metoden ble utviklet i 1957 av M. Meselson, F. Stahl og D. Winograd) er mye brukt for separasjon, isolering og analyse av individuelle komplementære DNA-tråder. For eksempel viste W. Shibalsky (USA), ved bruk av disse teknikkene for å separere DNA fra lambda-fagen, i 1967 - 1969 at begge fagkjedene er genetisk aktive , og ikke én, slik dette ble ansett for å være (S. Spiegelman, 1961). Det skal bemerkes at ideen om den genetiske betydningen av begge DNA-trådene til lambda-fagen for første gang ble uttrykt i USSR av SE Bresler (1961).
* (For sitt arbeid med genetikk til bakterier og virus ble A. Hershey sammen med M. Delbrück og S. Luria tildelt Nobelprisen i 1969.)
For å forstå organisasjonen og funksjonelle aktiviteten til genomet, er bestemmelsen av DNA-nukleotidsekvensen av største betydning. Søket etter metoder for slik bestemmelse utføres i mange laboratorier rundt om i verden. Siden slutten av 1950-tallet har M. Beer og hans samarbeidspartnere forsøkt å etablere DNA-sekvensen ved hjelp av elektronmikroskopi i USA, men så langt uten hell. På begynnelsen av 1950-tallet, fra de første verkene til Sinsheimer, Chargaff og andre forskere om enzymatisk nedbrytning av DNA, ble det kjent at forskjellige nukleotider i et DNA-molekyl er fordelt, men ikke tilfeldig, men ujevnt. Ifølge den engelske kjemikeren C. Barton (1961) konsentreres pyrimidiner (mer enn 70 %) hovedsakelig i form av de tilsvarende blokkene. A. L. Mazin og B. F. Vanyushin (1968 - 1969) fastslo at forskjellige DNA-er har ulik grad av pyrimidin-kohesjon og at i DNAet til dyreorganismer øker den markant når den beveger seg fra lavere til høyere. Dermed gjenspeiles utviklingen av organismer også i strukturen til deres genom. Det er derfor, for å forstå den evolusjonære prosessen som helhet, en sammenlignende studie av strukturen til nukleinsyrer er av spesiell betydning. Analyse av strukturen til biologisk viktige polymerer og først av alt DNA er ekstremt viktig for å løse mange spesielle problemer innen fylogenetikk og taksonomi.
Det er interessant å merke seg at den engelske fysiologen E. Lankester, som studerte hemoglobinene til bløtdyr, forutså ideene om molekylærbiologi for nøyaktig 100 år siden, skrev: «Kjemiske forskjeller mellom forskjellige arter og slekter av dyr og planter er like viktige for å avklare historien om deres opprinnelse som deres form. Hvis vi klart kunne fastslå forskjellene i organismenes molekylære organisering og funksjon, ville vi være i stand til å forstå opprinnelsen og utviklingen til forskjellige organismer mye bedre enn på grunnlag av morfologiske observasjoner " * . Betydningen av biokjemiske studier for taksonomi ble også understreket av V. L. Komarov, som skrev at "grunnlaget for alle til og med rent morfologiske karakterer, på grunnlag av hvilke vi klassifiserer og etablerer arter, er nettopp biokjemiske forskjeller" **.
* (E.R. Lankester. Uber das Vorcommen von Hemoglobin in den Muskeln der Mollusken und die Verbreitung desselben in den lebendigen Organismen.- "Pfluger" s Archiv fur die gesammte Physiol., 1871, Bd 4, 319.)
** (V. L. Komarov. Utvalgte verk, bind 1. M.-L., Publishing House of the Academy of Sciences of the USSR, 1945, s. 331.)
A. V. Blagoveshchenskii og S. L. Ivanov tok allerede på 1920-tallet de første skritt i vårt land for å belyse visse spørsmål om organismers evolusjon og systematikk på grunnlag av en sammenlignende analyse av deres biokjemiske sammensetning (se kapittel 2). Komparativ analyse av strukturen til proteiner og nukleinsyrer blir nå et stadig mer håndgripelig verktøy for taksonomer (se kapittel 21). Denne metoden for molekylærbiologi gjør det mulig ikke bare å avklare posisjonen til individuelle arter i systemet, men gjør det også nødvendig å ta en ny titt på selve prinsippene for klassifisering av organismer, og noen ganger å revidere hele systemet som helhet. , som skjedde for eksempel med systematikken til mikroorganismer. Utvilsomt, i fremtiden vil analysen av strukturen til genomet innta en sentral plass i kjemosystematikken til organismer.
Av stor betydning for utviklingen av molekylærbiologi var dechiffreringen av mekanismene for DNA-replikasjon og transkripsjon (se kapittel 24).
Proteinbiosyntese
Et viktig skifte i å løse problemet med proteinbiosyntese er assosiert med fremskritt i studiet av nukleinsyrer. I 1941 trakk T. Kasperson (Sverige) og i 1942 J. Brachet (Belgia) oppmerksomhet på det faktum at vev med aktiv proteinsyntese inneholder en økt mengde RNA. De konkluderte med at ribonukleinsyrer spiller en avgjørende rolle i proteinsyntesen. I 1953 ser det ut til at E. Gale og D. Fox har mottatt direkte bevis på direkte involvering av RNA i proteinbiosyntese: ifølge deres data undertrykte ribonuklease betydelig inkorporering av aminosyrer i bakteriecellelysater. Lignende data ble oppnådd av V. Olfri, M. Delhi og A. Mirsky (1953) på leverhomogenater. Senere avviste E. Gale sin korrekte idé om RNAs ledende rolle i proteinsyntese, og trodde feilaktig at aktiveringen av proteinsyntese i et cellefritt system skjedde under påvirkning av et annet stoff av ukjent natur. I 1954 fant P. Zamechnik, D. Littlefield, R. B. Khesin-Lurie og andre at den mest aktive inkorporeringen av aminosyrer skjer i RNA-rike fraksjoner av subcellulære partikler - mikrosomer. P. Zamechnik og E. Keller (1953 - 1954) fant at inkorporeringen av aminosyrer ble merkbart forbedret i nærvær av supernatanten under betingelser for ATP-regenerering. P. Sikevitz (1952) og M. Hoagland (1956) isolerte en proteinfraksjon (pH 5 fraksjon) fra supernatanten, som var ansvarlig for den skarpe stimuleringen av inkorporering av aminosyrer i mikrosomer. Sammen med proteiner ble det funnet en spesiell klasse RNA-er med lav molekylvekt, nå kalt overførings-RNA-er (tRNA-er), i supernatanten. I 1958 fant Hoagland og Zamechnik, samt P. Berg, R. Sweet og F. Allen og mange andre forskere ut at hver aminosyre krever sitt eget spesielle enzym, ATP og spesifikt tRNA, for å aktiveres. Det ble klart at tRNA-er utelukkende utfører funksjonen til adaptere, dvs. enheter som finner en plass på nukleinmatrisen (mRNA) for den tilsvarende aminosyren i det fremvoksende proteinmolekylet. Disse studiene bekreftet fullt ut adapterhypotesen til F. Crick (1957), som sørget for eksistensen av polynukleotidadaptere i cellen, som er nødvendige for riktig arrangement av aminosyrerester av det syntetiserte proteinet på nukleinmatrisen. Mye senere viste den franske vitenskapsmannen F. Chapville (1962) i laboratoriet til F. Lipman (Nobelprisen, 1953) i USA meget genialt og entydig at plasseringen av en aminosyre i et syntetisert proteinmolekyl er fullstendig bestemt av spesifikt tRNA som det er festet til. Cricks adapterhypotese ble utviklet av Hoagland og Zamechnik.
I 1958 ble følgende hovedstadier av proteinsyntese kjent: 1) aktivering av en aminosyre med et spesifikt enzym fra "pH 5-fraksjonen" i nærvær av ATP med dannelse av aminoacyladenylat; 2) binding av en aktivert aminosyre til et spesifikt tRNA med frigjøring av adenosinmonofosfat (AMP); 3) binding av aminoacyl-tRNA (tRNA lastet med en aminosyre) til mikrosomer og inkorporering av aminosyrer i et protein med tRNA-frigjøring. Hoagland (1958) bemerket at guanosintrifosfat (GTP) er nødvendig i det siste stadiet av proteinsyntesen.
Overføre RNA og gensyntese
Etter oppdagelsen av tRNA begynte aktive søk etter fraksjonering og bestemmelse av nukleotidsekvensen. Den amerikanske biokjemikeren R. Holly oppnådde størst suksess. I 1965 etablerte han strukturen til alanin tRNA fra gjær. Ved å bruke ribonukleaser (guanyl-RNase og pankreas-RNase) delte Holly nukleinsyremolekylet i flere fragmenter, bestemte nukleotidsekvensen i hver av dem separat, og rekonstruerte deretter sekvensen til hele alanin-tRNA-molekylet. Denne måten å analysere nukleotidsekvensen på kalles blokkmetoden. Hollys fortjeneste besto hovedsakelig i det faktum at han lærte å dele RNA-molekylet ikke bare i små biter, som mange gjorde før ham, men også i store fragmenter (kvardeler og halvdeler). Dette ga ham muligheten til å sette sammen individuelle småbiter korrekt og derved gjenskape den komplette nukleotidsekvensen til hele tRNA-molekylet (Nobelprisen, 1968).
Denne teknikken ble umiddelbart tatt i bruk av mange laboratorier rundt om i verden. I løpet av de neste to årene ble den primære strukturen til flere tRNA-er dechiffrert i USSR og i utlandet. A. A. Baev (1967) og medarbeidere etablerte nukleotidsekvensen i gjærvalin-tRNA for første gang. Til dags dato har mer enn et dusin forskjellige individuelle tRNA-er blitt studert. En særegen rekord i å bestemme nukleotidsekvensen ble satt i Cambridge av F. Senger og G. Brownlee. Disse forskerne utviklet en overraskende elegant metode for å separere oligonukleotider og sekvensere det såkalte 5 S (ribosomalt) RNA fra E. coli-celler (1968). Dette RNA består av 120 nukleotidrester og inneholder, i motsetning til tRNA, ikke ytterligere mindre baser, noe som i stor grad letter analysen av nukleotidsekvensen, og fungerer som unike landemerker for individuelle fragmenter av molekylet. For tiden, takket være bruken av metoden til Sanger og Brownlee, blir arbeidet med studiet av sekvensen til lange ribosomale RNA-er og noen virale RNA-er med suksess videreført i laboratoriet til J. Ebel (Frankrike) og andre forskere.
A. A. Baev og kollegaer (1967) fant at valin tRNA kuttet i to gjenoppretter sin makromolekylære struktur i løsning og, til tross for en defekt i primærstrukturen, har den funksjonelle aktiviteten til det opprinnelige (native) molekylet. Denne tilnærmingen - rekonstruksjonen av et kuttet makromolekyl etter fjerning av visse fragmenter - viste seg å være veldig lovende. Det er nå mye brukt for å belyse den funksjonelle rollen til individuelle seksjoner av visse tRNA-er.
De siste årene har det blitt oppnådd stor suksess med å oppnå krystallinske preparater av individuelle tRNAer. Mange tRNA-er har allerede blitt krystallisert i flere laboratorier i USA og England. Dette gjorde det mulig å studere strukturen til tRNA ved hjelp av røntgendiffraksjonsanalyse. I 1970 presenterte R. Bock de første røntgenmønstrene og tredimensjonale modellene av flere tRNA-er som han hadde laget ved University of Wisconsin. Disse modellene hjelper til med å bestemme lokaliseringen av individuelle funksjonelt aktive steder i tRNA og forstå de grunnleggende prinsippene for funksjonen til disse molekylene.
Dechiffreringen av naturen til den genetiske koden (se kapittel 24), som uten overdrivelse kan betraktes som den ledende prestasjonen for naturvitenskap på 1900-tallet, var av største betydning for å avsløre mekanismen for proteinsyntese og løse problemet spesifisiteten til denne prosessen.
R. Hollys oppdagelse av den primære strukturen til tRNA ga drivkraft til arbeidet til G. Korana * (USA) med syntese av oligonukleotider og rettet dem mot syntesen av en spesifikk biologisk struktur - et DNA-molekyl som koder for alanin-tRNA. De første trinnene i den kjemiske syntesen av korte oligonukleotider laget av Koranen for nesten 15 år siden kulminerte i 1970 med den første gensyntesen. Koran og hans samarbeidspartnere syntetiserte først kjemisk korte fragmenter av 8-12 nukleotidrester fra individuelle nukleotider. Disse fragmentene med en gitt nukleotidsekvens dannet spontant dobbelttrådete komplementære stykker med en overlapping på 4–5 nukleotider. Deretter ble disse ferdiglagde stykkene sammenføyd ende-til-ende i riktig rekkefølge ved hjelp av enzymet DNA-ligase. Således klarte Koranen, i motsetning til replikasjonen av DNA-molekyler, ifølge A. Kornberg ** (se kapittel 24), å gjenskape et naturlig dobbelttrådet DNA-molekyl etter et forhåndsplanlagt program iht. tRNA-sekvensen beskrevet av Holly. På samme måte pågår det nå arbeid med syntese av andre gener (M.N. Kolosov, Z.A. Shabarova, D.G. Knorre, 1970 - 1975).
* (For studiet av den genetiske koden ble G. Koran og M. Nirenberg tildelt Nobelprisen i 1968.)
** (For oppdagelsen av polymerase og DNA-syntese A. Kornberg, og for syntesen av RNA S. Ochoa i 1959 ble tildelt Nobelprisen.)
Mikrosomer, ribosomer, oversettelse
På midten av 1950-tallet ble det antatt at mikrosomer var sentrum for proteinsyntesen i cellen. Begrepet mikrosomer ble først introdusert i 1949 av A. Claude for å referere til fraksjonen av små granuler. Senere viste det seg at ikke hele fraksjonen av mikrosomer, bestående av membraner og granuler, men bare små ribonukleoproteinpartikler, er ansvarlig for proteinsyntesen. Disse partiklene ble i 1958 kalt ribosomer av R. Roberts.
Klassiske studier av bakterielle ribosomer ble utført av A. Tisier og J. Watson i 1958-1959. Bakterielle ribosomer viste seg å være noe mindre enn planter og dyr. J. Littleton (1960), M. Clark (1964) og E. N. Svetailo (1966) viste at ribosomene til kloroplastene til høyere planter og mitokondrier tilhører bakterietypen. A. Tisier og andre (1958) fant at ribosomer dissosieres i to ulike underenheter som inneholder ett RNA-molekyl hver. På slutten av 50-tallet ble det antatt at hvert ribosomalt RNA-molekyl består av flere korte fragmenter. Imidlertid var AS Spirin i 1960 den første som viste at RNA i underpartikler er representert av et kontinuerlig molekyl. D. Waller (1960), etter å ha separert ribosomale proteiner ved bruk av stivelsegelelektroforese, fant at de er svært heterogene. Til å begynne med tvilte mange på Wallers data, siden det så ut til at ribosomproteinet skulle være strengt homogent, som for eksempel TMV-proteinet. For tiden, som et resultat av forskningen til D. Waller, R. Trout, P. Traub og andre biokjemikere, har det blitt kjent at sammensetningen av de egentlige ribosompartiklene inkluderer mer enn 50 proteiner som er helt forskjellige i struktur. A. S. Spirin i 1963 var den første som utfoldet ribosomale subpartikler og viste at ribosomer er en kompakt vridd ribonukleoproteinstreng, som under visse forhold kan utfolde seg. I 1967 - 1968 M. Nomura rekonstruerte fullstendig en biologisk aktiv underenhet fra ribosomalt RNA og protein og oppnådde til og med ribosomer der protein og RNA tilhørte forskjellige mikroorganismer.
Rollen til ribosomalt RNA er fortsatt uklart. Det antas at det er den unike spesifikke matrisen som, under dannelsen av en ribosomal partikkel, hvert av de tallrike ribosomale proteinene finner et strengt definert sted (AS Spirin, 1968).
A. Rich (1962) oppdaget aggregater av flere ribosomer koblet sammen med en tråd av mRNA. Disse kompleksene ble kalt polysomer. Oppdagelsen av polysomer tillot Rich og Watson (1963) å foreslå at syntesen av polypeptidkjeden skjer på ribosomet, som så å si beveger seg langs mRNA-kjeden. Når ribosomet beveger seg langs mRNA-kjeden i partikkelen, leses informasjon ut og en proteinpolypeptidkjede dannes, og nye ribosomer fester seg vekselvis til den frigjorte leseenden av mRNA. Fra dataene til Rich og Watson, fulgte det at betydningen av polysomer i en celle ligger i masseproduksjonen av protein ved suksessiv lesing av matrisen av flere ribosomer samtidig.
Som et resultat av forskningen til M. Nirenberg, S. Ochoa, F. Lipman, G. Korana og andre i 1963 - 1970. det ble kjent at sammen med mRNA, ribosomer, ATP og aminoacyl-tRNA deltar et stort antall ulike faktorer i translasjonsprosessen, og selve translasjonsprosessen kan betinget deles inn i tre stadier - initiering, selve translasjonen og terminering.
Translasjonsinitiering betyr syntesen av den første peptidbindingen i det komplekse ribosomet - malpolynukleotid - aminoacyl-tRNA. Slik initierende aktivitet har ikke noe aminoacyl-tRNA, men av formylmetionyl-tRNA. Dette stoffet ble først isolert i 1964 av F. Senger og K. Marker. S. Bretcher og K. Marker (1966) viste at den initierende funksjonen til formylmetionyl-tRNA skyldes dets økte affinitet for peptidylsenteret i ribosomet. For starten av translasjonen er noen proteininitieringsfaktorer også ekstremt viktige, som ble isolert i laboratoriene til S. Ochoa, F. Gro og andre forskningssentre. Etter dannelsen av den første peptidbindingen i ribosomet, starter selve translasjonen, dvs. den sekvensielle tilsetningen av en aminoacylrest til C-terminalen av polypeptidet. Mange detaljer om oversettelsesprosessen ble studert av K. Monroe og J. Bishop (England), I. Rykhlik og F. Shorm (Tsjekkoslovakia), F. Lipman, M. Bretcher, W. Gilbert (USA) og andre forskere. I 1968 foreslo A. S. Spirin en original hypotese for å forklare mekanismen til ribosomet. Drivmekanismen som sikrer alle romlige bevegelser av tRNA og mRNA under translasjon er den periodiske åpningen og lukkingen av ribosomsubpartikler. Oversettelsestermineringen er kodet i selve den lesbare matrisen, som inneholder termineringskodonene. Som vist av S. Brenner (1965 - 1967), er trillinger UAA, UAG og UGA slike kodoner. M. Capecci (1967) identifiserte også spesielle proteintermineringsfaktorer. AS Spirin og LP Gavrilova beskrev den såkalte "ikke-enzymatiske" proteinsyntesen i ribosomer (1972 - 1975) uten deltagelse av proteinfaktorer. Denne oppdagelsen er viktig for å forstå opprinnelsen og utviklingen av proteinbiosyntese.
Regulering av gen- og proteinaktivitet
Etter problemet med spesifisiteten til proteinsyntese, viste problemet med regulering av proteinsyntese, eller, det samme, regulering av genaktivitet, seg å være i første rekke i molekylærbiologien.
Den funksjonelle ulikheten til celler og undertrykkelsen og aktiveringen av gener assosiert med den har lenge tiltrukket seg oppmerksomheten til genetikere, men inntil nylig forble den virkelige mekanismen for å kontrollere genaktivitet ukjent.
De første forsøkene på å forklare den regulatoriske aktiviteten til gener var assosiert med studiet av histonproteiner. Til og med Steadman-ektefellene * på begynnelsen av 40-tallet av XX-tallet. antydet at det er histoner som kan spille hovedrollen i dette fenomenet. Deretter fikk de de første klare dataene om forskjeller i den kjemiske naturen til histonproteiner. For tiden øker antallet fakta som vitner til fordel for denne hypotesen hvert år.
* (E. Stedman, E. Stedman. De grunnleggende proteinene til cellekjerner.- Filosof. Trans. Roy. soc. London, 1951, v. 235, 565 - 595.)
Samtidig samler det seg en økende mengde data, noe som indikerer at reguleringen av genaktivitet er en mye mer kompleks prosess enn den enkle interaksjonen mellom genseksjoner med histonproteinmolekyler. I 1960 - 1962 i laboratoriet til RB Khesin-Lurie, ble det funnet at faggener begynner å leses ikke-samtidig: T2 faggener kan deles inn i tidlige, hvis funksjon skjedde i de første minuttene av infeksjon av en bakteriecelle, og sene, som begynte å syntetisere mRNA etter fullføringen av arbeidet med tidlige gener.
I 1961 foreslo de franske biokjemikerne F. Jacob og J. Monod et opplegg for regulering av genaktivitet, som spilte en eksepsjonell rolle i forståelsen av cellens reguleringsmekanismer generelt. I henhold til skjemaet til Jacob og Monod, i tillegg til strukturelle (informasjons-) gener, inneholder DNA også gener-regulatorer og gener-operatører. Regulatorgenet koder for syntesen av et spesifikt stoff - en repressor, som kan feste seg både til induktoren og til operatørgenet. Operatørgenet er knyttet til strukturelle gener, mens regulatorgenet er lokalisert i en viss avstand fra dem. Hvis det ikke er noen induktor i miljøet, for eksempel laktose, binder repressoren syntetisert av regulatorgenet seg til operatørgenet og blokkerer det, og slår av arbeidet til hele operonet (en blokk av strukturelle gener sammen med operatøren) som styrer dem). Enzymdannelse skjer ikke under disse forholdene. Hvis en induktor (laktose) vises i mediet, binder produktet av regulatorgenet, repressoren, seg til laktose og fjerner blokken fra operatørgenet. I dette tilfellet blir arbeidet med det strukturelle genet som koder for syntesen av enzymet mulig, og enzymet (laktose) vises i mediet.
I følge Jacob og Monod er dette reguleringsskjemaet anvendelig for alle adaptive enzymer og kan finne sted både under undertrykkelse, når dannelsen av enzymet undertrykkes av et overskudd av reaksjonsproduktet, og under induksjon, når introduksjonen av et substrat forårsaker syntesen av enzymet. For studier av regulering av genaktivitet ble Jacob og Monod tildelt Nobelprisen i 1965.
I utgangspunktet virket denne ordningen for langsøkt. Senere viste det seg imidlertid at reguleringen av gener i henhold til dette prinsippet ikke bare skjer i bakterier, men også i andre organismer.
Siden 1960 har en fremtredende plass i molekylærbiologi blitt okkupert av studier av organiseringen av genomet og strukturen til kromatin i eukaryote organismer (J. Bonner, R. Britten, W. Olfrey, P. Walker, Yu. S. Chentsov , IB Zbarsky og andre .) og regulering av transkripsjon (A. Mirsky, G. P. Georgiev, M. Bernstiel, D. Goll, R. Tsanev, R. I. Salganik). I lang tid forble undertrykkerens natur ukjent og kontroversiell. I 1968 viste M. Ptashne (USA) at et protein er en repressor. Han isolerte det i laboratoriet til J. Watson og fant ut at repressoren virkelig har en affinitet for induseren (laktose) og samtidig "gjenkjenner" operatorgenet til lac-operonet og binder seg spesifikt til det.
I løpet av de siste 5 - 7 årene har det blitt innhentet data om tilstedeværelsen av en annen kontrollcelle med genaktivitet - promoteren. Det viste seg at i nærheten av operatørstedet, som produktet syntetisert på genregulatoren - proteinsubstansen til repressoren, er festet til, er det et annet nettsted som også bør tilskrives medlemmene av reguleringssystemet. av genaktivitet. Et proteinmolekyl av enzymet RNA-polymerase er festet til dette stedet. I promotorregionen må gjensidig gjenkjennelse av den unike nukleotidsekvensen i DNA og den spesifikke konfigurasjonen av RNA-polymeraseproteinet skje. Implementeringen av prosessen med å lese genetisk informasjon med en gitt sekvens av gener av operonet ved siden av promoteren vil avhenge av gjenkjenningseffektiviteten.
I tillegg til ordningen beskrevet av Jacob og Monod, er det andre mekanismer for genregulering i cellen. F. Jacob og S. Brenner (1963) fastslo at reguleringen av bakteriell DNA-replikasjon styres på en bestemt måte av cellemembranen. Eksperimentene til Jacob (1954) på induksjon av forskjellige profeter viste overbevisende at under påvirkning av forskjellige mutagene faktorer i cellen til lysogene bakterier, begynner selektiv replikasjon av profaggenet, og replikering av vertsgenomet blokkeres. I 1970 rapporterte F. Bell at små DNA-molekyler kan passere fra kjernen inn i cytoplasmaet og bli transkribert der.
Dermed kan genaktivitet reguleres på nivået av replikasjon, transkripsjon og translasjon.
Det er gjort betydelige fremskritt i å studere reguleringen av ikke bare syntesen av enzymer, men også deres aktivitet. A. Novik og L. Szilard påpekte fenomenene med regulering av aktiviteten til enzymer i cellen tilbake på 1950-tallet. G. Umbarger (1956) fant at i cellen er det en svært rasjonell måte å undertrykke aktiviteten til enzymet ved sluttproduktet av tilbakemeldingskjeden av reaksjoner. Som etablert av J. Monod, J. Change, F. Jacob, A. Purdy og andre forskere (1956 - 1960), kan reguleringen av enzymaktivitet utføres i henhold til det allosteriske prinsippet. Enzymet eller en av dets underenheter, i tillegg til affinitet for substratet, har en affinitet for et av produktene i reaksjonskjeden. Under påvirkning av et slikt signalprodukt endrer enzymet sin konformasjon på en slik måte at det mister aktivitet. Som et resultat blir hele kjeden av enzymatiske reaksjoner slått av helt i begynnelsen. D. Wiman og R. Woodward (1952; Nobelprisvinner, 1965) påpekte den essensielle rollen til proteinkonformasjonsendringer i enzymatiske reaksjoner, og i en viss forstand tilstedeværelsen av en allosterisk effekt.
Struktur og funksjon av proteiner
Som et resultat av arbeidet til T. Osborn, G. Hofmeister, A. Gurber, F. Schulz og mange andre på slutten av 1800-tallet. Mange animalske og vegetabilske proteiner er oppnådd i krystallinsk form. Omtrent samtidig ble molekylvektene til visse proteiner bestemt ved hjelp av forskjellige fysiske metoder. Så i 1891 rapporterte A. Sabaneev og N. Alexandrov at molekylvekten til ovalbumin er 14 000; i 1905 fant E. Reid at molekylvekten til hemoglobin er 48 000. Den polymere strukturen til proteiner ble oppdaget i 1871 av G. Glasivetz og D. Gaberman. Ideen om en peptidbinding av individuelle aminosyrerester i proteiner ble fremmet av T. Curtius (1883). Arbeid med kjemisk kondensering av aminosyrer (E. Schaal, 1871; G. Schiff, 1897; L. Balbiano og D. Traschiatti, 1900) og syntesen av heteropolypeptider (E. Fisher, 1902 - 1907, Nobelprisen, 1902) førte til utviklingen av de grunnleggende prinsippene den kjemiske strukturen til proteiner.
Det første krystallinske enzymet (urease) ble oppnådd i 1926 av J. Sumner (Nobelprisen, 1946), og i 1930 oppnådde J. Northrop (Nobelprisen, 1946) krystallinsk pepsin. Etter disse arbeidene ble det klart at enzymer er av proteinkarakter. I 1940 isolerte M. Kunits krystallinsk RNase. I 1958 var mer enn 100 krystallinske enzymer og over 500 ikke-krystallinske enzymer allerede kjent. Å skaffe høyt rensede preparater av individuelle proteiner bidro til dechiffreringen av deres primære struktur og makromolekylære organisering.
Av stor betydning for utviklingen av molekylærbiologi generelt og menneskelig genetikk, spesielt, var oppdagelsen av L. Pauling (1940) av unormalt hemoglobin S, isolert fra erytrocyttene til mennesker med en alvorlig arvelig sykdom, sigdcelleanemi. I 1955 - 1957 W. Ingram brukte "fingeravtrykk"-metoden utviklet av F. Sanger (flekker dannet av individuelle peptider under kromatografi på papir) for å analysere produktene av hydrolyse av hemoglobin S med alkali og trypsin. I 1961 rapporterte Ingram at hemoglobin S skiller seg fra normalt hemoglobin bare i naturen til én aminosyrerest: i normalt hemoglobin er en glutaminsyrerest i den syvende posisjonen i kjeden, og i hemoglobin S, en valinrest. Dermed ble Paulings antakelse (1949) om at sigdcelleanemi er en sykdom av molekylær natur fullstendig bekreftet. En arvelig endring i bare én aminosyrerest i hver halvdel av hemoglobinmakromolekylet fører til at hemoglobin mister sin evne til å løse seg lett opp ved lav oksygenkonsentrasjon og begynner å krystallisere, noe som fører til forstyrrelse av cellestrukturen. Disse studiene viste tydelig at strukturen til et protein er en strengt definert aminosyresekvens som er kodet i genomet. Arbeidene til K. Anfinsen (1951) vitnet om den eksepsjonelle betydningen av den primære strukturen til et protein i dannelsen av en unik biologisk aktiv konformasjon av et makromolekyl. Anfinsen viste at den biologisk aktive makrostrukturen til bukspyttkjertelribonuklease, som går tapt som et resultat av restaurering, er forhåndsbestemt av aminosyresekvensen og kan dukke opp spontant under oksidasjon av SH-grupper av cysteinrester med dannelse av disulfid-tverrbindinger i strengt definerte steder i peptidkjeden til enzymet.
Til dags dato har virkningsmekanismen til et stort antall enzymer blitt studert i detalj og strukturen til mange proteiner er bestemt.
I 1953 etablerte F. Sanger aminosyresekvensen til insulin. : Dette proteinet består av to polypeptidkjeder forbundet med to disulfid-tverrbindinger. En av kjedene inneholder kun 21 aminosyrerester, mens den andre inneholder 30 rester. Sanger brukte omtrent 10 år på å tyde strukturen til dette relativt enkle proteinet. I 1958 ble han tildelt Nobelprisen for denne fremragende forskningen. Etter opprettelsen av V. Stein og S. Moore (1957) av en automatisk analysator av aminosyrer, akselererte identifiseringen av produkter av delvis hydrolyse av proteiner betydelig. I 1960 rapporterte Stein og Moore det allerede. at de var i stand til å bestemme sekvensen til ribonuklease, hvis peptidkjede er representert av 124 aminosyrerester. Samme år, i laboratoriet til G. Schramm i Tübingen (Tyskland), bestemte F. Anderer og andre aminosyresekvensen i TMV-proteinet. Deretter ble aminosyresekvensen bestemt i myoglobin (A. Edmunson) og α- og β-kjeder av humant hemoglobin (G. Braunitzer, E. Schroeder, etc.), lysozym fra eggprotein (J. Jollet, D. Keyfield) . I 1963 etablerte F. Shorm og B. Keil (Tsjekkoslovakia) aminosyresekvensen i chymotrypsinogen-molekylet. Samme år ble aminosyresekvensen til trypsinogen bestemt (F. Shorm, D. Walsh). I 1965 etablerte K. Takahashi den primære strukturen til ribonuklease T1. Deretter ble aminosyresekvensen bestemt for flere proteiner.
Som kjent er det endelige beviset på riktigheten av definisjonen av en bestemt struktur syntesen. I 1969 var R. Merifield (USA) den første som utførte kjemisk syntese av bukspyttkjertelribonuklease. Ved hjelp av sin metode for syntese på en fastfasebærer, tilførte Merifield den ene aminosyren etter den andre til kjeden i samsvar med sekvensen som ble beskrevet av Stein og Moore. Som et resultat fikk han et protein som i kvalitet var identisk med bukspyttkjertelribonuklease A. For oppdagelsen av strukturen til ribonuklease ble V. Stein, S. Moore og K. Anfinsen tildelt Nobelprisen i 1972. Denne naturlige proteinsyntesen åpner for enorme muligheter, og peker på muligheten for å lage proteiner i samsvar med en forhåndsplanlagt sekvens.
Fra røntgenstrukturstudier av W. Astbury (1933) fulgte det at peptidkjedene til proteinmolekyler er vridd eller stablet på en strengt definert måte. Siden den gang har mange forfattere uttrykt ulike hypoteser om hvordan proteinkjeder brettes, men frem til 1951 forble alle modeller spekulative konstruksjoner som ikke samsvarte med eksperimentelle data. I 1951 publiserte L. Pauling og R. Corey en serie strålende artikler der teorien om den sekundære strukturen til proteiner, teorien om α-helixen, til slutt ble formulert. Sammen med dette ble det også kjent at proteiner også har en tertiær struktur: α-helixen til peptidkjeden kan foldes på en bestemt måte, og danner en ganske kompakt struktur.
I 1957 foreslo J. Kendrew og hans medarbeidere først en tredimensjonal modell av strukturen til myoglobin. Denne modellen ble deretter foredlet over flere år, inntil det endelige arbeidet dukket opp i 1961 med en karakterisering av den romlige strukturen til dette proteinet. I 1959 etablerte M. Perutz og kolleger den tredimensjonale strukturen til hemoglobin. Forskere brukte mer enn 20 år på dette arbeidet (de første røntgenstrålene av hemoglobin ble oppnådd av Perutz i 1937). Siden hemoglobinmolekylet består av fire underenheter, etter å ha dechiffrert organisasjonen, beskrev Perutz derved først den kvaternære strukturen til proteinet. For sitt arbeid med å bestemme den tredimensjonale strukturen til proteiner, ble Kendrew og Perutz tildelt Nobelprisen i 1962.
Opprettelsen av en romlig modell av strukturen til hemoglobin av Perutz TILLATT. å komme nærmere forståelsen av funksjonsmekanismen til dette proteinet, som som kjent utfører oksygentransport i dyreceller. Tilbake i 1937 kom F. Gaurowitz til den konklusjon at samspillet mellom hemoglobin og oksygen, luft skulle være ledsaget av en endring i strukturen til proteinet. På 1960-tallet oppdaget Perutz og medarbeidere et merkbart skifte i hemoglobinkjedene etter oksidasjonen, forårsaket av forskyvning av jernatomer som et resultat av binding med oksygen. På dette grunnlaget ble ideer om "pusten" av proteinmakromolekyler dannet.
I 1960 begynte D. Phillips og hans samarbeidspartnere røntgendiffraksjonsstudier av lysozymmolekylet. I 1967 var de mer eller mindre i stand til å etablere detaljene for organiseringen av dette proteinet og lokaliseringen av individuelle atomer i molekylet. I tillegg fant Phillips ut arten av tilsetningen av lysozym til substratet (triacetylglukosamin). Dette gjorde det mulig å gjenskape mekanismen til dette enzymet. Kunnskap om den primære strukturen og makromolekylære organisasjonen gjorde det derfor mulig ikke bare å etablere naturen til de aktive sentrene til mange enzymer, men også å fullt ut avsløre funksjonsmekanismen til disse makromolekylene.
Bruken av elektronmikroskopimetoder bidro til å avsløre prinsippene for den makromolekylære organiseringen av slike komplekse proteinformasjoner som kollagen, fibrinogen, kontraktile muskelfibriller osv. På slutten av 1950-tallet ble det foreslått modeller av det muskulære kontraktile apparatet. Av eksepsjonell betydning for å forstå mekanismen for muskelkontraksjon var oppdagelsen av V. A. Engelgardt og M. N. Lyubimova (1939) av ATPase-aktiviteten til myosin. Dette betydde at muskelkontraksjonshandlingen er basert på en endring i de fysisk-kjemiske egenskapene og makromolekylær organisering av det kontraktile proteinet under påvirkning av adenosintrifosforsyre (se også kapittel 11).
Virologisk forskning har vært avgjørende for å forstå prinsippene for å sette sammen biologiske strukturer (se kapittel 25).
Uløste problemer
De viktigste fremskrittene innen moderne molekylærbiologi har blitt oppnådd hovedsakelig som et resultat av studiet av nukleinsyrer. Men selv på dette området er langt fra alle problemer løst. Spesielt vil det kreves stor innsats for å dechiffrere hele nukleotidsekvensen til genomet. Dette problemet er på sin side uløselig knyttet til problemet med DNA-heterogenitet og krever utvikling av nye avanserte metoder for fraksjonering og isolering av individuelle molekyler fra cellens totale genetiske materiale.
Inntil nå har innsatsen hovedsakelig vært fokusert på separat studie av proteiner og nukleinsyrer. I cellen er disse biopolymerene uløselig knyttet til hverandre og fungerer hovedsakelig i form av nukleoproteiner. Derfor er behovet for å studere interaksjonen mellom proteiner og nukleinsyrer nå blitt spesielt akutt. Problemet med gjenkjennelse av visse deler av nukleinsyrer av proteiner blir aktualisert. Det er allerede skissert trinn for å studere en slik interaksjon mellom disse biopolymerene, uten hvilken en fullstendig forståelse av strukturen og funksjonene til kromosomer, ribosomer og andre strukturer er utenkelig. Uten dette er det også umulig å forstå reguleringen av genaktivitet og til slutt dechiffrere prinsippene for arbeidet med proteinsyntesemekanismer. Etter arbeidet til Jacob og Monod dukket det opp noen nye data om den regulatoriske betydningen av membraner i syntesen av kjernefysisk materiale. Dette utgjør problemet med en dypere studie av rollen til membraner i reguleringen av DNA-replikasjon. Generelt har problemet med regulering av genaktivitet og celleaktivitet generelt blitt et av de viktigste problemene i moderne molekylærbiologi.
Den nåværende tilstanden til biofysikk
I nær sammenheng med molekylærbiologiens problemer fortsatte utviklingen av biofysikk. Interessen for dette området av biologi ble stimulert, på den ene siden, av behovet for en omfattende studie av effekten av ulike typer stråling på kroppen, og på den andre siden av behovet for å studere den fysiske og fysiske -kjemiske grunnlaget for livsfenomener som oppstår på molekylært nivå.
Å få nøyaktig informasjon om molekylære strukturer og prosessene som foregår i dem ble mulig som et resultat av bruken av nye fine fysiske og kjemiske metoder. Basert på oppnåelsene av elektrokjemi var det mulig å forbedre metoden for å måle bioelektriske potensialer ved å bruke ioneselektive elektroder (G. Eisenman, B.P. Nikolsky, Khuri, 50-60-årene). I økende grad kommer infrarød spektroskopi (med bruk av laserenheter) i praksis, noe som gjør det mulig å studere konformasjonsendringene i proteiner (I. Plotnikov, 1940). Verdifull informasjon er også gitt av metoden for elektronparamagnetisk resonans (E.K. Zavoisky, 1944) og den biokjemiluminescerende metoden (B.N. Tarusov et al., 1960), som gjør det spesielt mulig å bedømme transporten av elektroner under oksidative prosesser.
På 1950-tallet var biofysikk allerede i ferd med å få en sterk posisjon. Det er behov for å utdanne kvalifiserte spesialister. Hvis i 1911 i Europa bare University of Pécs, i Ungarn, hadde en leder for biofysikk, så eksisterer slike stoler i 1973 i nesten alle store universiteter.
I 1960 ble International Society of Biophysicists organisert. I august 1961 fant den første internasjonale biofysiske kongressen sted i Stockholm. Den andre kongressen ble holdt i 1965 i Paris, den tredje - i 1969 i Boston, den fjerde - i 1972 i Moskva.
I biofysikk er det et klart skille mellom to områder med ulikt innhold – molekylær biofysikk og cellulær biofysikk. Denne utmerkelsen får også et organisatorisk uttrykk: separate avdelinger for disse to områdene av biofysikk blir opprettet. Ved Moskva-universitetet ble den første avdelingen for biofysikk opprettet i 1953 ved Fakultetet for biologi og jordvitenskap, og litt senere dukket Institutt for biofysikk opp ved Det fysiske fakultet. Avdelinger ble organisert etter samme prinsipp ved mange andre universiteter.
Molekylær biofysikk
De siste årene har sammenhengen mellom molekylær biofysikk og molekylærbiologi blitt stadig sterkere, og nå er det noen ganger vanskelig å fastslå hvor skillelinjen mellom dem går. I et generelt angrep på problemet med arvelig informasjon er et slikt samarbeid mellom biofysikk og molekylærbiologi uunngåelig.
Hovedretningen i forskningsarbeidet er studiet av nukleinsyrenes fysikk – DNA og RNA. Bruken av metodene ovenfor og fremfor alt røntgendiffraksjonsanalyse bidro til dechiffreringen av molekylstrukturen til nukleinsyrer. For tiden pågår intensiv forskning for å studere oppførselen til disse syrene i løsninger. Spesiell oppmerksomhet rettes mot konformasjonsovergangene "helix-coil", som studeres av endringer i viskositet, optiske og elektriske parametere. I forbindelse med studiet av mekanismene for mutagenese, utvikles det studier for å studere effekten av ioniserende stråling på oppførselen til nukleinsyrer i løsninger, samt effekten av stråling på nukleinsyrene til virus og fager. Effekten av ultrafiolett stråling, hvorav noen spektrale områder er kjent for å bli godt absorbert av nukleinsyrer, ble gjenstand for en omfattende analyse. En stor andel av denne typen forskning er påvisning av aktive radikaler av nukleinsyrer og proteiner ved hjelp av elektronparamagnetisk resonans. Med bruken av denne metoden er fremveksten av en hel uavhengig retning assosiert.
Problemet med å kode DNA- og RNA-informasjon og dens overføring under proteinsyntese har lenge vært av interesse for molekylær biofysikk, og fysikere har gjentatte ganger uttrykt visse betraktninger om dette emnet (E. Schrödinger, G. Gamow). Dechiffreringen av den genetiske koden forårsaket en rekke teoretiske og eksperimentelle studier på strukturen til DNA-spiralen, mekanismen for glidning og vridning av trådene, og studiet av de fysiske kreftene involvert i disse prosessene.
Molekylær biofysikk gir betydelig hjelp til molekylærbiologi i å studere strukturen til proteinmolekyler ved hjelp av røntgendiffraksjonsanalyse, som først ble brukt i 1930 av J. Bernal. Det var som et resultat av bruken av fysiske metoder i kombinasjon med biokjemiske (enzymatiske metoder) at den molekylære konformasjonen og sekvensen av aminosyrer i en rekke proteiner ble avslørt.
Moderne elektronmikroskopiske studier, som har avslørt tilstedeværelsen av komplekse membransystemer i celler og dens organeller, har stimulert forsøk på å forstå deres molekylære struktur (se kapittel 10 og 11). Den kjemiske sammensetningen av membraner og spesielt egenskapene til lipidene deres studeres in vivo. Det ble funnet at sistnevnte er i stand til overoksidasjon og ikke-enzymatiske reaksjoner av kjedeoksidasjon (Yu. A. Vladimirov og F. F. Litvin, 1959; B. N. Tarusov et al., 1960; I. I. Ivanov, 1967), noe som fører til membrandysfunksjon. For å studere sammensetningen av membraner begynte man også å bruke metoder for matematisk modellering (V. Ts. Presman, 1964 - 1968; M. M. Shemyakin, 1967; Yu. A. Ovchinnikov, 1972).
Cellulær biofysikk
En betydelig begivenhet i biofysikkens historie var dannelsen på 1950-tallet av klare ideer om termodynamikken til biologiske prosesser, som et resultat av hvilke antakelser om muligheten for uavhengig generering av energi i levende celler, i strid med termodynamikkens andre lov. , forsvant til slutt. Å forstå virkemåten til denne loven i biologiske systemer er forbundet med introduksjonen av den belgiske forskeren I. Prigogine (1945) * i biologisk termodynamikk av konseptet åpne systemer som utveksler energi og materie med det ytre miljøet. Prigogine viste at positiv entropi dannes i levende celler under arbeidsprosesser i samsvar med termodynamikkens andre lov. Ligningene han introduserte bestemte forholdene under hvilke den såkalte stasjonære tilstanden oppstår (tidligere ble den også kalt dynamisk likevekt), der mengden fri energi (negentropi) som kommer inn i cellene med mat kompenserer for forbruket, og positiv entropi er produksjon. Denne oppdagelsen forsterket den generelle biologiske ideen om den uatskillelige forbindelsen mellom det ytre og indre miljøet til celler. Det markerte begynnelsen på en virkelig studie av termodynamikken til levende systemer, inkludert modelleringsmetoden (A. Burton, 1939; A. G. Pasynsky, 1967).
* (Den generelle teorien om åpne systemer ble først fremmet av L. Bertalanffy i 1932.)
I henhold til det grunnleggende prinsippet for biotermodynamikk er en nødvendig betingelse for eksistensen av liv stasjonaritet i utviklingen av dets biokjemiske prosesser, for gjennomføringen av disse er det nødvendig å koordinere hastighetene til en rekke metabolske reaksjoner. På bakgrunn av den nye biofysiske termodynamikken har det oppstått en trend som skiller ut ytre og indre faktorer som sikrer denne koordineringen av reaksjoner og gjør den stabil. I løpet av de siste to tiårene har en stor rolle i å opprettholde den stasjonære tilstanden til systemet av inhibitorer og spesielt antioksidanter blitt avslørt (B.N. Tarusov og A.I. Zhuravlev, 1954, 1958). Det er fastslått at påliteligheten til stasjonær utvikling er assosiert med miljøfaktorer (temperatur) og de fysisk-kjemiske egenskapene til cellemiljøet.
Moderne prinsipper for biotermodynamikk har gjort det mulig å gi en fysisk-kjemisk tolkning av tilpasningsmekanismen. I følge våre data kan tilpasning til miljøforhold bare skje hvis kroppen, når de endrer seg, er i stand til å etablere stasjonaritet i utviklingen av biokjemiske reaksjoner (B.N. Tarusov, 1974). Spørsmålet oppsto om å utvikle nye metoder som ville tillate å vurdere den stasjonære tilstanden in vivo og forutsi mulige brudd. Innføringen av kybernetiske prinsipper for selvregulerende systemer i biotermodynamikk og forskning på prosessene for biologisk tilpasning lover stor fordel. Det ble klart at for å løse problemet med stabiliteten til den stasjonære tilstanden, er det viktig å ta hensyn til de såkalte forstyrrende faktorene, som spesielt inkluderer ikke-enzymatiske reaksjoner av lipidoksidasjon. Nylig har studier av prosessene med overoksidasjon i lipidfasene til levende celler og veksten av aktive radikale produkter som bryter med membranens regulerende funksjoner utvidet seg. Kilden til informasjon om disse prosessene er både påvisning av aktive peroksidradikaler og peroksidforbindelser av biolipider (A. Tappel, 1965; I.I. Ivanov, 1965; E.B. Burlakova, 1967 og andre). For å oppdage radikaler brukes biokjemiluminescens, som oppstår i lipidene til levende celler under rekombinasjonen.
På grunnlag av fysisk-kjemiske ideer om stabiliteten til steady state, oppsto biofysiske ideer om tilpasning av planter til endringer i miljøforhold som et brudd på hemmende antioksidantsystemer (B.N. Tarusov, Ya. E. Doskoch, B. M. Kitlaev, A. M. Agaverdiev , 1968–1972). Dette åpnet for muligheten for å vurdere egenskaper som frostbestandighet og salttoleranse, samt å komme med passende spådommer i valg av landbruksplanter.
På 1950-tallet ble en ultrasvak glød oppdaget - biokjemiluminescens av en rekke biologiske objekter i de synlige og infrarøde delene av spekteret (B. N. Tarusov, A. I. Zhuravlev, A. I. Polivoda). Dette ble mulig som et resultat av utviklingen av metoder for registrering av supersvake lysstrømmer ved bruk av fotomultiplikatorer (L. A. Kubetsky, 1934). Som et resultat av biokjemiske reaksjoner som oppstår i en levende celle, gjør biokjemiluminescens det mulig å bedømme viktige oksidative prosesser i elektronoverføringskjedene mellom enzymer. Oppdagelsen og studiet av biokjemiluminescens er av stor teoretisk og praktisk betydning. Så, BN Tarusov og Yu. B. Kudryashov bemerker den store rollen til produktene av oksidasjon av umettede fettsyrer i mekanismen for forekomsten av patologiske tilstander som utvikler seg under påvirkning av ioniserende stråling, i karsinogenese og andre brudd på de normale funksjonene av cellen.
På 1950-tallet, i forbindelse med den raske utviklingen av kjernefysikk, dukket radiobiologi, som studerer den biologiske effekten av ioniserende stråling, opp fra biofysikken. Produksjonen av kunstige radioaktive isotoper, etableringen av termonukleære våpen, atomreaktorer og utviklingen av andre former for praktisk bruk av atomenergi har med all sin akutthet ført til problemet med å beskytte organismer mot de skadelige effektene av ioniserende stråling, og utvikle teoretiske grunnlag for forebygging og behandling av strålesyke. For å gjøre dette var det først og fremst nødvendig å finne ut hvilke komponenter i cellen og koblinger av metabolisme som er de mest sårbare.
Studieobjektet i biofysikk og radiobiologi var å belyse arten av de primære kjemiske reaksjonene som oppstår i levende underlag under påvirkning av strålingsenergi. Her var det viktig ikke bare å forstå mekanismene til dette fenomenet, men også å kunne påvirke prosessen med å bytte fysisk energi mot kjemisk energi, for å redusere dens koeffisient for "nyttig" virkning. Arbeid i denne retningen ble initiert av studiene ved skolen til N. N. Semenov (1933) i USSR og D. Hinshelwood (1935) i England.
En viktig plass i radiobiologisk forskning ble okkupert av studiet av graden av strålingsmotstand til forskjellige organismer. Det ble funnet at økt radioresistens (for eksempel hos ørkengnagere) skyldes den høye antioksidantaktiviteten til cellemembranlipider (M. Chang et al., 1964; N. K. Ogryzov et al., 1969). Det viste seg at tokoferoler, vitamin K og tioforbindelser spiller en viktig rolle i dannelsen av antioksidantegenskapene til disse systemene (II Ivanov et al., 1972). De siste årene har studier av mekanismene for mutagenese også vakt mye oppmerksomhet. For dette formålet studeres effekten av ioniserende stråling på oppførselen til nukleinsyrer og proteiner in vitro, så vel som i virus og fager (A. Gustafson, 1945 - 1950).
Kampen for en ytterligere økning i effektiviteten av kjemisk beskyttelse, søket etter mer effektive hemmere og prinsipper for inhibering er fortsatt biofysikkens hovedoppgaver i denne retningen.
Det er gjort fremskritt i studiet av eksiterte tilstander av biopolymerer, som bestemmer deres høye kjemiske aktivitet. Den mest vellykkede var studiet av eksiterte tilstander som oppstår på det primære stadiet av fotobiologiske prosesser - fotosyntese og syn.
Dermed er det gitt et solid bidrag til å forstå den primære aktiveringen av molekylene i plantepigmentsystemer. Den store betydningen av overføring (migrering) av energien til eksiterte tilstander uten tap fra aktiverte pigmenter til andre substrater er fastslått. En stor rolle i utviklingen av disse ideene ble spilt av de teoretiske verkene til A. N. Terenin (1947 og senere). A. A. Krasnovsky (1949) oppdaget og studerte reaksjonen av reversibel fotokjemisk reduksjon av klorofyll og dets analoger. Det er nå en generell oppfatning at det i nær fremtid vil være mulig å reprodusere fotosyntese under kunstige forhold (se også kapittel 5).
Biofysikere fortsetter å jobbe med å avdekke naturen til muskelkontraksjon og mekanismene for nerveeksitasjon og -ledning (se kapittel 11). Forskning på mekanismene for overgangen fra en opphisset tilstand til en normal tilstand har også blitt av aktuell betydning. Den eksiterte tilstanden anses nå som et resultat av en autokatalytisk reaksjon, og inhibering anses som en konsekvens av en skarp mobilisering av hemmende antioksidantaktivitet som et resultat av molekylære omorganiseringer i forbindelser som tokoferol (II Ivanov, OR Kols, 1966; OR Kols, 1970).
Det viktigste generelle problemet med biofysikk er fortsatt kunnskapen om de kvalitative fysiske og kjemiske egenskapene til levende materie. Egenskaper som levende biopolymerers evne til selektivt å binde kalium eller polarisere elektrisk strøm kan ikke bevares selv med den mest forsiktige fjerning fra kroppen. Derfor fortsetter cellulær biofysikk å intensivt utvikle kriterier og metoder for livstidsstudier av levende materie.
Til tross for ungdommen innen molekylærbiologi, er fremgangen den har gjort på dette området virkelig fantastisk. På relativt kort tid har genets natur og de grunnleggende prinsippene for organisering, reproduksjon og funksjon blitt etablert. Dessuten har ikke bare in vitro-reproduksjon av gener blitt utført, men også for første gang er den fullstendige syntesen av selve genet fullført. Den genetiske koden er fullstendig dechiffrert og det viktigste biologiske problemet med spesifisiteten til proteinbiosyntese er løst. De viktigste måtene og mekanismene for proteindannelse i cellen er identifisert og studert. Den primære strukturen til mange transport-RNA, spesifikke adaptermolekyler som oversetter språket til nukleinmaler til språket til aminosyresekvensen til det syntetiserte proteinet, er fullstendig bestemt. Aminosyresekvensen til mange proteiner er fullstendig dechiffrert og den romlige strukturen til noen av dem er etablert. Dette gjorde det mulig å belyse prinsippet og detaljene for funksjonen til enzymmolekyler. Den kjemiske syntesen av et av enzymene, ribonuklease, ble utført. De grunnleggende prinsippene for organisering av ulike subcellulære partikler, mange virus og fager er etablert, og de viktigste måtene for deres biogenese i cellen har blitt avdekket. Tilnærminger for å forstå måtene for regulering av genaktivitet og belysning av de regulatoriske mekanismene for vital aktivitet har blitt oppdaget. Allerede en enkel liste over disse funnene indikerer at andre halvdel av det 20. århundre. var preget av enorme fremskritt innen biologi, noe som først og fremst skyldes en dybdestudie av strukturen og funksjonene til biologisk viktige makromolekyler – nukleinsyrer og proteiner.
Prestasjoner innen molekylærbiologi blir allerede brukt i praksis i dag og gir konkrete resultater innen medisin, landbruk og enkelte bransjer. Det er ingen tvil om at avkastningen til denne vitenskapen vil øke hver dag. Imidlertid bør hovedresultatet fortsatt vurderes at under påvirkning av suksessene til molekylærbiologi, har tilliten til eksistensen av ubegrensede muligheter på veien til å avsløre livets hemmeligste hemmeligheter styrket seg.
I fremtiden vil tilsynelatende nye måter å studere den biologiske formen for materiens bevegelse åpnes på - biologien vil bevege seg fra det molekylære nivået til det atomære nivået. Men nå er det sannsynligvis ikke en eneste forsker som realistisk kan forutsi utviklingen av molekylærbiologi selv for de neste 20 årene.
Fremskritt i studiet av nukleinsyrer og proteinbiosyntese har ført til opprettelsen av en rekke metoder av stor praktisk betydning innen medisin, landbruk og en rekke andre industrier.
Etter at den genetiske koden og de grunnleggende prinsippene for lagring og implementering av arvelig informasjon ble studert, stoppet utviklingen av molekylærbiologi, siden det ikke fantes noen metoder som gjorde det mulig å manipulere gener, isolere og endre dem. Fremveksten av disse metodene skjedde på 1970-1980-tallet. Dette ga en kraftig drivkraft til utviklingen av dette vitenskapsfeltet, som fortsatt blomstrer i dag. Først og fremst dreier disse metodene seg om å oppnå individuelle gener og deres introduksjon i celler fra andre organismer (molekylær kloning og transgenese, PCR), samt metoder for å bestemme nukleotidsekvensen i gener (DNA- og RNA-sekvensering). Disse metodene vil bli diskutert mer detaljert nedenfor. Vi starter med den enkleste grunnleggende metoden, elektroforese, og går så videre til mer komplekse metoder.
DNA ELEKTROFORESE
Det er den grunnleggende metoden for å jobbe med DNA, som brukes sammen med nesten alle andre metoder for å isolere de ønskede molekylene og analysere resultatene. Gelelektroforese brukes til å skille DNA-fragmenter etter lengde. DNA er en syre, dens molekyler inneholder fosforsyrerester, som spalter et proton og får en negativ ladning (fig. 1).
Derfor, i et elektrisk felt, beveger DNA-molekyler seg mot anoden - en positivt ladet elektrode. Dette skjer i en elektrolyttløsning som inneholder ladningsbærerioner, på grunn av hvilken denne løsningen leder strøm. For å skille fragmentene brukes en tett gel laget av polymerer (agarose eller polyakrylamid). DNA-molekyler "vikler seg" i det jo mer, jo lengre de er, og derfor beveger de lengste molekylene seg tregest, og kortest - raskest (fig. 2). Før eller etter elektroforese behandles gelen med fargestoffer som binder seg til DNA og fluorescerer i ultrafiolett lys, og man får et mønster av bånd i gelen (se fig. 3). For å bestemme lengden av DNA-fragmenter i en prøve, sammenlignes de med en markør, dvs. et sett med fragmenter av standardlengder avsatt parallelt på samme gel (fig. 4).



De viktigste verktøyene for å arbeide med DNA er enzymer som utfører DNA-transformasjoner i levende celler: DNA-polymeraser, DNA-ligaser og restriksjonsendonukleaser, eller restriksjonsenzymer. DNA-polymerase DNA-templatesyntese utføres, som gjør at DNA kan forplantes i et reagensrør. DNA-ligaser sy sammen DNA-molekyler eller helbrede hullene i dem. Restriksjonsendonukleaser, eller restriksjoner, kutt DNA-molekyler i henhold til strengt definerte sekvenser, som lar deg kutte ut individuelle fragmenter fra den totale massen av DNA. Disse fragmentene kan i noen tilfeller inneholde individuelle gener.
restriksjoner
Sekvenser gjenkjent av restriksjonsenzymer er symmetriske, og brudd kan oppstå midt i en slik sekvens eller med et skifte (på samme sted i begge DNA-trådene). Virkningsskjemaet for forskjellige typer restriktaser er vist i fig. 1. I det første tilfellet oppnås de såkalte "stumpe" endene, og i det andre - "klebrige" ender. Ved "klebrige" ender av bunnen er kjeden kortere enn den andre, det dannes en enkeltstrenget seksjon med en symmetrisk sekvens som er lik i begge ender.
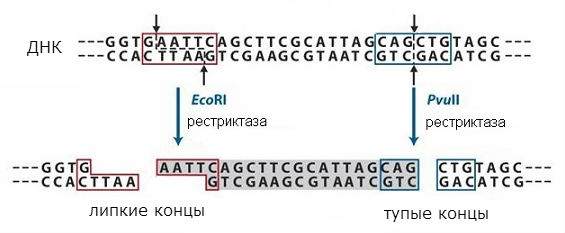
Endesekvensene vil være de samme når et hvilket som helst DNA spaltes med et gitt restriksjonsenzym og kan slås sammen igjen fordi de har komplementære sekvenser. De kan ligeres med DNA-ligase for å danne et enkelt molekyl. Dermed er det mulig å kombinere fragmenter av to forskjellige DNA og få den såkalte rekombinant DNA. Denne tilnærmingen brukes i metoden for molekylær kloning, som gjør det mulig å oppnå individuelle gener og introdusere dem i celler som kan danne proteinet kodet i genet.
molekylær kloning
Molekylær kloning bruker to DNA-molekyler - et innlegg som inneholder genet av interesse, og vektor- DNA fungerer som en bærer. Innsatsen "sys" inn i vektoren ved hjelp av enzymer, og får et nytt, rekombinant DNA-molekyl, deretter introduseres dette molekylet i vertsceller, og disse cellene danner kolonier på et næringsmedium. En koloni er et avkom av én celle, dvs. en klon, alle celler i kolonien er genetisk identiske og inneholder samme rekombinante DNA. Derav begrepet "molekylær kloning", det vil si å oppnå en klon av celler som inneholder et DNA-fragment av interesse for oss. Etter at koloniene som inneholder innsatsen av interesse for oss er oppnådd, er det mulig å karakterisere denne innsatsen ved forskjellige metoder, for eksempel for å bestemme dens nøyaktige rekkefølge. Cellene kan også produsere proteinet kodet av innskuddet hvis det inneholder et funksjonelt gen.
Når et rekombinant molekyl introduseres i celler, skjer den genetiske transformasjonen av disse cellene. Transformasjon- prosessen med absorpsjon av en celle av en organisme av et fritt DNA-molekyl fra miljøet og dets integrering i genomet, noe som fører til opptreden i en slik celle av nye arvelige egenskaper for den, karakteristiske for organisme-donoren av DNA . For eksempel, hvis det innsatte molekylet inneholder et gen for resistens mot antibiotikumet ampicillin, vil de transformerte bakteriene vokse i dets nærvær. Før transformasjon forårsaket ampicillin deres død, det vil si at et nytt tegn vises i de transformerte cellene.
VEKTORER
En vektor må ha en rekke egenskaper:
For det første er det et relativt lite DNA-molekyl som enkelt kan manipuleres.
For det andre, for at DNA skal bevares og reproduseres i en celle, må det inneholde en bestemt sekvens som sikrer replikasjonen (opprinnelsen til replikasjon, eller replikasjonsopprinnelsen).
For det tredje må den inneholde markørgen, som sikrer seleksjon av kun de cellene som vektoren har gått inn i. Vanligvis er dette antibiotikaresistensgener - da i nærvær av et antibiotikum dør alle celler som ikke inneholder vektoren.
Genkloning utføres oftest i bakterieceller, da de er enkle å dyrke og formere seg raskt. I en bakteriecelle er det vanligvis ett stort sirkulært DNA-molekyl, flere millioner basepar langt, som inneholder alle genene som er nødvendige for bakterier - det bakterielle kromosomet. I tillegg til det er det i noen bakterier små (flere tusen basepar) sirkulært DNA, kalt plasmider(Fig. 2). De, i likhet med hoved-DNAet, inneholder en nukleotidsekvens som gir DNA'ets evne til å replikere (ori). Plasmider replikerer uavhengig av hoved-(kromosomalt) DNA, derfor er de tilstede i cellen i et stort antall kopier. Mange av disse plasmidene bærer antibiotikaresistensgener, som gjør det mulig å skille celler som bærer plasmidet fra normale celler. Mer vanlig brukes plasmider som bærer to gener som gir resistens mot to antibiotika, som tetracyklin og amycilin. Det finnes enkle metoder for å isolere slikt plasmid-DNA fritt fra DNA-et til bakteriens hovedkromosom.

BETYDNING AV TRANSGENESIS
Overføring av gener fra en organisme til en annen kalles transgenese, og slike modifiserte organismer - transgenisk. Metoden for genoverføring til mikrobielle celler brukes til å oppnå rekombinante proteinpreparater for medisin, spesielt humane proteiner som ikke forårsaker immunavvisning - interferoner, insulin og andre proteinhormoner, cellevekstfaktorer, samt proteiner for produksjon av vaksiner. I mer komplekse tilfeller, når proteinmodifisering bare forløper riktig i eukaryote celler, brukes transgene cellekulturer eller transgene dyr, spesielt husdyr (først og fremst geiter), som skiller ut de nødvendige proteinene til melk, eller proteiner isoleres fra blodet deres. Slik oppnås antistoffer, blodkoagulasjonsfaktorer og andre proteiner. Transgenesemetoden gir kulturplanter som er resistente mot ugressmidler og skadedyr og har andre nyttige egenskaper. Ved å bruke transgene mikroorganismer for å rense avløpsvann og bekjempe forurensning, er det til og med transgene mikrober som kan bryte ned olje. I tillegg er transgene teknologier uunnværlige i vitenskapelig forskning – utviklingen av biologi i dag er utenkelig uten rutinemessig bruk av genmodifikasjons- og overføringsmetoder.
molekylær kloningsteknologi
setter inn
For å få et individuelt gen fra en hvilken som helst organisme, blir alt kromosomalt DNA isolert fra det og spaltet med ett eller to restriksjonsenzymer. Enzymer velges slik at de ikke kutter genet av interesse for oss, men gjør brudd langs kantene, og i plasmid-DNA gjør de ett brudd i et av resistensgenene, for eksempel mot ampicillin.
Den molekylære kloningsprosessen inkluderer følgende trinn:
Klipp og sy - konstruksjon av et enkelt rekombinant molekyl fra en innsats og en vektor.
Transformasjon er introduksjonen av et rekombinant molekyl i celler.
Seleksjon - utvalg av celler som mottok en vektor med en innsetting.
kutting og sying
Plasmid-DNA behandles med de samme restriksjonsenzymene, og det blir til et lineært molekyl hvis det velges et slikt restriksjonsenzym som introduserer 1 brudd i plasmidet. Som et resultat vises de samme klebrige endene i endene av alle de resulterende DNA-fragmentene. Når temperaturen senkes, slutter disse endene seg tilfeldig og ligeres med DNA-ligase (se fig. 3).

En blanding av sirkulære DNA-er med forskjellig sammensetning oppnås: noen av dem vil inneholde en viss DNA-sekvens av kromosomalt DNA koblet til bakterielt DNA, andre vil inneholde fragmenter av kromosomalt DNA koblet sammen, og atter andre vil inneholde et redusert sirkulært plasmid eller dets dimer (Fig. 4).

transformasjon
Deretter utføres denne blandingen genetisk transformasjon bakterier som ikke inneholder plasmider. Transformasjon- prosessen med absorpsjon av en celle av en organisme av et fritt DNA-molekyl fra miljøet og dets integrering i genomet, noe som fører til opptreden i en slik celle av nye arvelige egenskaper for den, karakteristiske for organisme-donoren av DNA . Bare ett plasmid kan gå inn og formere seg i hver celle. Slike celler plasseres på et fast næringsmedium som inneholder antibiotikumet tetracyklin. Celler som ikke fikk plasmidet vil ikke vokse på dette mediet, og cellene som bærer plasmidet danner kolonier, som hver inneholder etterkommere av kun én celle, dvs. alle celler i en koloni bærer det samme plasmidet (se fig. 5).

Utvalg
Deretter er oppgaven å isolere bare cellene som vektoren med innskuddet har gått inn i, og å skille dem fra celler som bare bærer vektoren uten innskuddet eller som ikke bærer vektoren i det hele tatt. Denne prosessen med å velge de riktige cellene kalles utvalg. For dette, søk selektive markører- vanligvis antibiotikaresistensgener i vektoren, og selektive medier som inneholder antibiotika eller andre selektive stoffer.
I eksemplet vi vurderer, blir celler fra kolonier dyrket i nærvær av ampicillin subkulturert på to medier: den første inneholder ampicillin, og den andre inneholder tetracyklin. Kolonier som kun inneholder plasmidet vil vokse på begge medier, mens kolonier som inneholder innsatt kromosomalt DNA i plasmidene ikke vil vokse på mediet med tetracyklin (fig. 5). Blant dem, ved spesielle metoder, velges de som inneholder genet av interesse for oss, dyrkes i tilstrekkelige mengder, og plasmid-DNA isoleres. Fra det, ved å bruke de samme restriktasene som ble brukt for å oppnå rekombinant DNA, kuttes det individuelle genet av interesse ut. DNAet til dette genet kan brukes til å bestemme sekvensen av nukleotider, introdusere i en hvilken som helst organisme for å oppnå nye egenskaper eller syntetisere det ønskede proteinet. Denne metoden for genisolering kalles molekylær kloning.
FLUORESCENTE PROTEINER
Det er veldig praktisk å bruke fluorescerende proteiner som markørgener i studier av eukaryote organismer. Genet for det første fluorescerende proteinet, grønt fluorescerende protein (GFP) ble isolert fra maneten Aqeuorea victoria og introdusert i ulike modellorganismer (se fig. 6) I 2008 mottok O. Shimomura, M. Chalfi og R. Tsien Nobelprisen for oppdagelsen og anvendelsen av dette proteinet.

Deretter ble genene for andre fluorescerende proteiner - rødt, blått, gult - isolert. Disse genene har blitt kunstig modifisert for å produsere proteiner med de ønskede egenskapene. Mangfoldet av fluorescerende proteiner er vist i fig. 7, som viser en petriskål med bakterier som inneholder gener for ulike fluorescerende proteiner.

påføring av fluorescerende proteiner
Det fluorescerende proteingenet kan smeltes sammen med genet til et hvilket som helst annet protein, og under translasjon vil det dannes et enkelt protein - et translasjonsfusjonsprotein, eller fusjon(fusjonsprotein), som fluorescerer. Dermed er det mulig å studere for eksempel lokaliseringen (plasseringen) av eventuelle proteiner av interesse i cellen, deres bevegelse. Ved å bruke ekspresjonen av fluorescerende proteiner kun i visse typer celler, er det mulig å merke celler av disse typene i en flercellet organisme (se fig. 8 - musehjerne, hvor individuelle nevroner har forskjellige farger på grunn av en viss kombinasjon av fluorescerende proteiner gener). Fluorescerende proteiner er et uunnværlig verktøy i moderne molekylærbiologi.

PCR
En annen metode for å skaffe gener kalles polymerasekjedereaksjon (PCR). Den er basert på evnen til DNA-polymeraser til å fullføre den andre DNA-strengen langs den komplementære strengen, slik det skjer i celler under DNA-replikasjon.
Opprinnelsen til replikasjon i denne metoden er gitt av to små biter av DNA kalt frø, eller primere. Disse primerne er komplementære til endene av genet av interesse på to DNA-tråder. Først blandes det kromosomale DNA som genet skal isoleres fra med frø og varmes opp til 99 ° C. Dette fører til brudd av hydrogenbindinger og divergens av DNA-tråder. Etter det senkes temperaturen til 50-70 ca C (avhengig av lengden og rekkefølgen av frø). Under disse forholdene blir primerne festet til komplementære regioner av kromosomalt DNA, og danner en vanlig dobbelhelix (se fig. 9). Deretter tilsettes en blanding av alle fire nukleotidene som trengs for DNA-syntese og DNA-polymerase. Enzymet forlenger primerne ved å bygge dobbelttrådet DNA fra festepunktet til primerne, dvs. fra endene av et gen til enden av et enkeltstrenget kromosommolekyl. 
Hvis blandingen nå varmes opp igjen, vil de kromosomale og nysyntetiserte kjedene spres. Etter avkjøling vil frø igjen slutte seg til dem, som tas i stort overskudd (se fig. 10).

På de nylig syntetiserte kjedene vil de slutte seg ikke til den enden som den første syntesen startet fra, men til den motsatte, siden DNA-kjedene er antiparallelle. Derfor, i den andre syntesesyklusen, vil bare sekvensen som tilsvarer genet bli fullført på slike kjeder (se fig. 11).

Denne metoden bruker DNA-polymerase fra termofile bakterier som tåler koking og fungerer ved temperaturer på 70-80 ° C, det trenger ikke tilsettes hver gang, men det er nok å legge det til i begynnelsen av eksperimentet. Ved å gjenta oppvarmings- og avkjølingsprosedyrene i samme sekvens, kan vi doble antall sekvenser i hver syklus, avgrenset i begge ender av de innførte frøene (se fig. 12).

Etter omtrent 25 slike sykluser vil antallet kopier av genet øke med mer enn en million ganger. Slike mengder kan lett separeres fra kromosomalt DNA som føres inn i reagensrøret og brukes til ulike formål.
DNA-sekvensering
En annen viktig prestasjon er utviklingen av metoder for å bestemme sekvensen av nukleotider i DNA - DNA-sekvensering(fra engelsk sekvens - sekvens). For å gjøre dette er det nødvendig å skaffe gener rene fra annet DNA ved å bruke en av de beskrevne metodene. Deretter separeres DNA-kjedene ved oppvarming og en primer merket med radioaktivt fosfor eller en fluorescerende markør tilsettes dem. Vær oppmerksom på at ett frø er tatt, komplementært til en kjede. Deretter tilsettes DNA-polymerase og en blanding av 4 nukleotider. En slik blanding deles i 4 deler og ett av nukleotidene tilsettes hver, modifisert slik at den ikke inneholder en hydroksylgruppe på det tredje atomet av deoksyribose. Hvis et slikt nukleotid er inkludert i den syntetiserte DNA-kjeden, vil dets forlengelse ikke kunne fortsette, fordi polymerase vil ikke ha noe sted å feste neste nukleotid. Derfor blir DNA-syntese etter inkludering av et slikt nukleotid avbrutt. Disse nukleotidene, kalt dideoksynukleotider, tilsettes mye mindre enn vanlig, så kjedeterminering skjer bare av og til og i hver kjede på forskjellige steder. Resultatet er en blanding av kjeder av forskjellig lengde, hver med samme nukleotid på slutten. Dermed tilsvarer kjedelengden nukleotidnummeret i sekvensen som studeres, for eksempel hvis vi hadde et adenyldideoksynukleotid, og de resulterende kjedene var 2, 7 og 12 nukleotider lange, så var adenin i andre, syvende og tolvte posisjon i genet. Den resulterende blandingen av kjeder kan enkelt separeres etter størrelse ved hjelp av elektroforese, og de syntetiserte kjedene kan identifiseres ved radioaktivitet på røntgenfilm (se fig. 10).

Det viser seg bildet vist nederst på bildet, kalt radioautograf. Når vi beveger oss langs den fra bunn til topp og leser bokstaven over kolonnene i hver sone, får vi nukleotidsekvensen vist i figuren til høyre for autografen. Det viste seg at syntesen ikke bare stoppes av dideoksynukleotider, men også av nukleotider der en eller annen kjemisk gruppe, for eksempel et fluorescerende fargestoff, er festet til sukkerets tredje posisjon. Hvis hvert nukleotid er merket med sitt eget fargestoff, vil sonene som oppnås ved å separere de syntetiserte kjedene lyse med et annet lys. Dette gjør det mulig å utføre reaksjonen i ett reagensrør samtidig for alle nukleotider og ved å separere de resulterende kjedene etter lengde, å identifisere nukleotidene etter farge (se fig. 11).

Slike metoder gjorde det mulig å bestemme sekvensene ikke bare til individuelle gener, men også å lese hele genomer. Enda raskere metoder for å bestemme nukleotidsekvenser i gener er nå utviklet. Hvis det første menneskelige genomet ble dekodet av et stort internasjonalt konsortium ved bruk av den første gitte metoden om 12 år, kan det andre, ved bruk av det andre, på tre år, nå gjøres dette på en måned. Dette lar deg forutsi en persons disposisjon for mange sykdommer og iverksette tiltak på forhånd for å unngå dem.
31.2
For venner!
referanse
Molekylærbiologi vokste ut av biokjemi i april 1953. Utseendet er assosiert med navnene til James Watson og Francis Crick, som oppdaget strukturen til DNA-molekylet. Oppdagelsen ble gjort mulig gjennom studiet av genetikk, bakterier og biokjemien til virus. Yrket som molekylærbiolog er ikke utbredt, men i dag er dens rolle i det moderne samfunnet veldig stor. Et stort antall sykdommer, inkludert de som manifesteres på genetisk nivå, krever at forskere finner løsninger på dette problemet.
Beskrivelse av aktivitet
Virus og bakterier muterer hele tiden, noe som betyr at medisiner ikke lenger hjelper en person og sykdommer blir uhåndterlige. Molekylærbiologiens oppgave er å komme i forkant av denne prosessen og utvikle en ny kur mot sykdommer. Forskere jobber i henhold til et veletablert opplegg: blokkerer årsaken til sykdommen, eliminerer arvelighetsmekanismene og lindrer dermed pasientens tilstand. Det finnes en rekke sentre, klinikker og sykehus rundt om i verden hvor molekylærbiologer utvikler nye behandlinger for å hjelpe pasienter.
Job ansvar
Ansvaret til en molekylærbiolog inkluderer studiet av prosesser inne i cellen (for eksempel endringer i DNA under utviklingen av svulster). Eksperter studerer også egenskapene til DNA, deres effekt på hele organismen og en enkelt celle. Slike studier utføres for eksempel på grunnlag av PCR (polymerasekjedereaksjon), som lar deg analysere kroppen for infeksjoner, arvelige sykdommer og bestemme biologisk forhold.
Funksjoner ved karrierevekst
Yrket som molekylærbiolog er ganske lovende på sitt felt og hevder allerede i dag å være det første i rangeringen av fremtidens medisinske profesjoner. En molekylærbiolog trenger forresten ikke å være i dette feltet hele tiden. Dersom det er et ønske om å bytte yrke, kan han omskolere seg til salgsleder for laboratorieutstyr, begynne å utvikle instrumenter for ulike studier, eller åpne egen virksomhet.